Az élő anyagot felépítő molekulák egyik fontos funkciója, hogy az életfolyamatokkal kapcsolatos kémiai reakciókat speciális katalizátorként felgyorsítsák (enzimek). E molekulák másik fontos funkcióként információt tárolnak és hordoznak (nukleinsavak). Emellett az élőlények ezeket a makromolekulákat képesek önállóan előállítani és sokszorosítani (replikáció).
Az élő anyag speciális szerveződési és vezérlési viszonyainak kialakulására vonatkozó egyik hipotézist Manfred Eigen állította fel. Szerinte mind az élő anyag, mind a vezérlést végző információhordozó molekulák megjelenésében és önszerveződésében a ciklikus folyamatok jelentették a legfontosabb előrelépést, és a ciklus tagjai alapján a fejlődést fázisokra tagolhatjuk. Az első fontos fázis a biokémiai reakciók körfolyamattá alakulása volt, azaz olyan reakciók alakultak ki, amelyekben a folyamat végterméke szolgáltatta az egész körfolyamat kiindulási molekuláját (24 ábra A: A,B,C,X = molekulák). A második fázist olyan körfolyamatok kialakulása jelentette, amelyekben a körfolyamat tagjaiként nem szubsztrát- vagy termékmolekulák, hanem reakciókat katalizáló enzimek foglaltak helyet (24. ábra B: E1,E2,E3,En = enzimek). A harmadik fázisban pedig megjelentek azok az információhordozó molekulák, amelyek saját maguk replikációját is képesek voltak irányítani (autokatalitikus önreplikáció), és ők alkották a körfolyamat egyes komponenseit (24. ábra C: I1,I2,I3,In = önreplikálódó információhordozó molekulák).
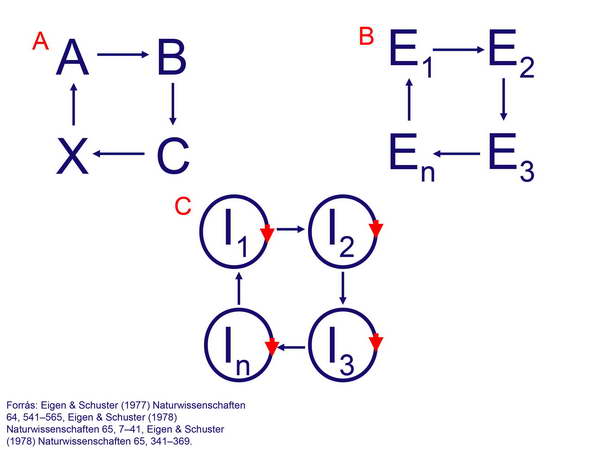
24. ábra
Később ezek a ciklusok megfelelő hierarchikus rendet alakítottak ki, és Eigen nyomán az autokatalitikus képességű vezérlőmolekulákból és az általuk felépített katalizátorokból felépülő ciklusokat hiperciklusoknak nevezzük (25. ábra):
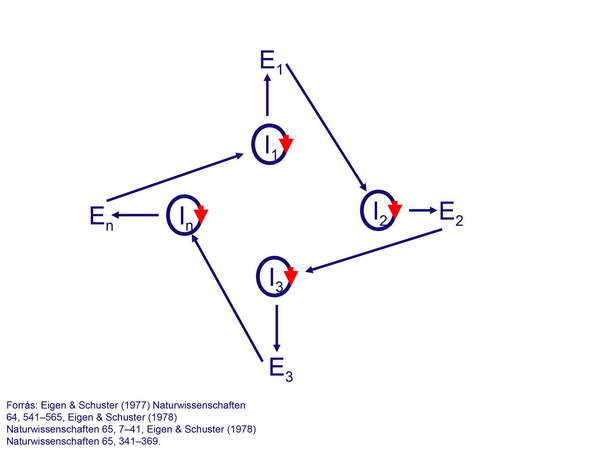
25. ábra
Ezek megjelenése tette lehetővé az információhordozó makromolekulák kettős működését (információ-átadás és vezérlés), ilyenkor ugyanis az önreplikációra képes információhordozó molekulák információ-tartalma alapján fognak elkészülni azok a katalitikus funkciót végző egységek (enzimek), amelyek aztán a megfelelő biokémiai reakciókat megvalósítják.
A Nobel-díjas Francis Crick híres dogmája szerint a biológiai rendszerekben az információátadás iránya: DNS-ből az RNS felé, és onnan a fehérje felé mutat (centrális dogma). Ez az elképzelés még az 1960-as években keletkezett, közvetlenül a genetikai kód megfejtését követően. Ez a teória az első komolyabb kihívást akkor kapta, amikor felfedezték a retrovírusokat, amelyek RNS-templát alapján képesek DNS-t szintetizálni. De logikai alapon is több támadás érte a centrális dogmát, hiszen a sejtekben mind a DNS, mind az RNS szintézisét fehérjék végzik, azaz a Crick szerinti DNS-----RNS-----fehérje sorrend valójában nem egy lineáris egyirányú lánc, hanem egy önmagába visszatérő kör, azaz ciklus. Felmerült tehát a szokásos „tyúk vagy tojás” dilemma, azaz az a kérdés, hogy ennek az önmagába visszatérő ciklusnak mi volt a kezdőpontja. Más megfogalmazásban: hogyan ment végbe az információ-hordozás és vezérlés evolúciója. A kérdés még nem kapott egyértelmű választ, de az egyik legelfogadottabb hipotézis szerint az első élő rendszer megjelenésekor az információ hordozója az RNS volt, mely tárolta is a genetikai információt és irányította is a primitív sejtek működését.
Az RNS-molekula ősi jellegét több kétségtelen tény támasztja alá.
Az RNS-molekula egyszálú, amely mindenképpen egy ősi jelleg a kétszálú DNS-sel szemben.
Az egyszálú RNS képes saját kópiáját 2 egymást követő lépésben létrehozni úgy, hogy először az eredeti szál szekvenciája szerepel templátként, ami alapján felépül annak komplementer szálja. A második lépésben ez a komplementer szál szerepel templátként, ami alapján felépül annak komplementere, azaz az eredeti szekvencia (26. ábra).

26. ábra
Hasonló funkcióra a DNS önmagában képtelen.
Az RNS-molekulában található ribóz létrejöhet abiotikus úton is, pl. formaldehidből (HCOH), amely viszont már jelen volt a Föld ősi anyagai között. Ezzel ellentétben a DNS-ben előforduló dezoxiribóz a ribózból alakul ki minden ma élő sejtben egy enzim (tehát fehérje) által katalizált reakcióban. Mindezek igen erős bizonyítékok annak alátámasztására, hogy az RNS megjelenése feltehetőleg megelőzte a DNS megjelenését. Más megfontolások alapján (a fehérjék szintéziséhez RNS-re van szükség), az is kimondható, hogy az RNS megjelenése a fehérjék megjelenését is meg kellett, hogy előzze.
Talán a legdöntőbb bizonyíték az RNS korai szerepére az RNS katalitikus funkciójának felfedezése (ribozim) volt 1982-ben. (27. ábra)
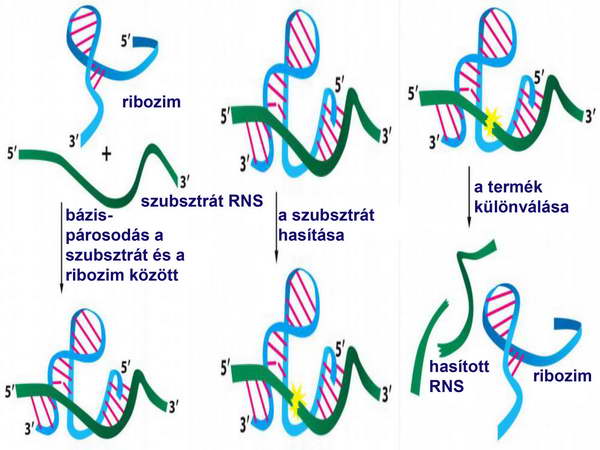
27. ábra
Ez annál is inkább fontos, mert mind a mai napig hasonló funkciót a DNS-nél nem írtak le. Feltételezések szerint ez a katalitikus funkció azt is lehetővé tette, hogy az RNS –molekula irányítani tudja saját maga replikációját (28. ábra).
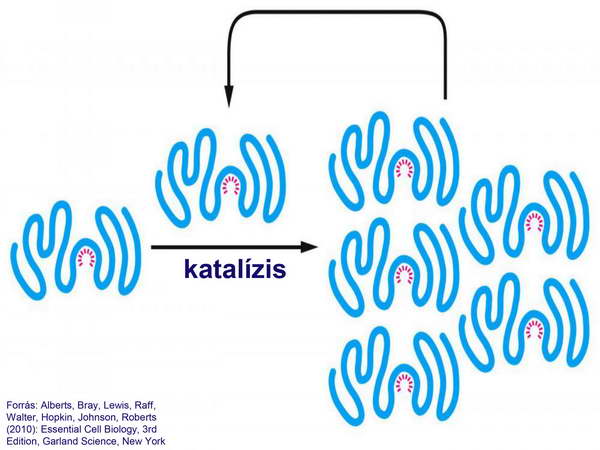
28. ábra
Mindezek alapján tehát úgy tűnik, hogy az első sejtek számára az RNS jelentette és egyben egyesítette a genetikai, strukturális és katalitikus funkciókat. Kezdetben az RNS katalizálhatta saját maga szintézisét, és a katalitikus funkciókat is ő maga töltötte be. Később jöhettek létre az RNS nukleotid-sorrendje alapján felépülő katalizátorok (fehérjék), és csak a harmadik lépésben vette át az irányítást a DNS (29. ábra).
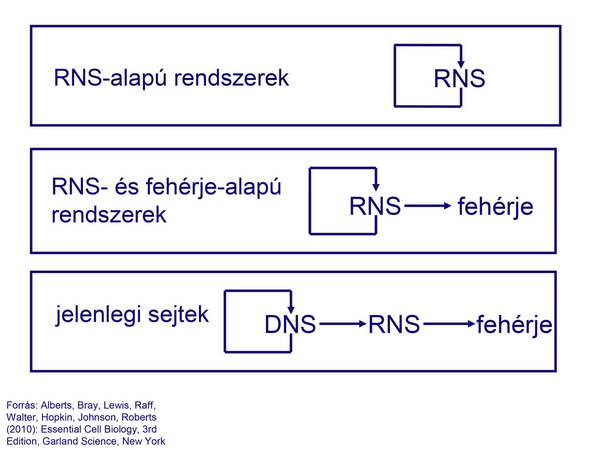
29. ábra
A fejlődés első (RNS-en alapuló) korszakát hívják „RNS-világ”-nak is (30. ábra).
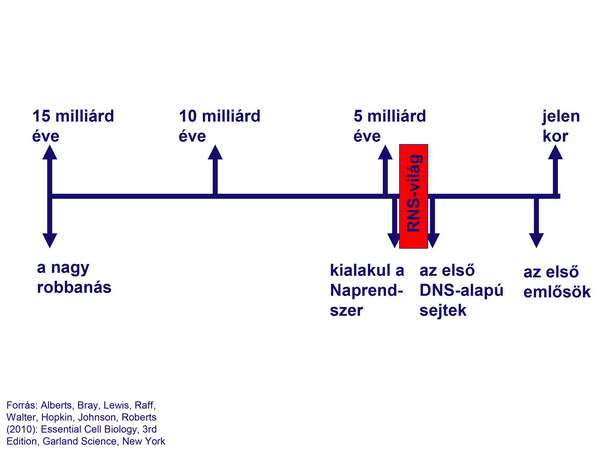
30. ábra