Az első sejtek prokarióták voltak, amelyek nevüket az elkülönült sejtmag hiányáról kapták. Emellett azonban számos igen fontos tulajdonság jellemzi őket, amelyek sokkal fontosabbak, bár valószínüleg semmilyen kapcsolatban sincsenek a valódi sejtmag hiányával. Ilyenek:
Kis méret (maximum néhány μm).
A belső állomány egyszerű, strukturálatlan, homogén jellegű.
A DNS (kromoszóma) cirkuláris, amely viszont több kópiában is előfordulhat, sőt esetenként „extrakromoszómális” DNS is található, lásd plazmidok.
A DNS nem tekeredik hisztonokra, nincs kromatinállomány.
Korábban úgy gondolták, hogy a prokariótákban egyáltalán nincs citoszkeletális rendszer, de újabban leírtak olyan struktúrákat, amelyek az eukarióták citoszkeletális rendszerével mutatnak hasonlóságot. Ilyenek: FtsZ, MreB, crescentin.
Jellemző a sejtfal jelenléte, amely felépítését tekintve általában peptidoglikán (az Archaeákban nem peptidoglikán, hanem fehérje, sőt a sejtfal el is maradhat).
A citoplazmában 70S típusú riboszómák találhatók.
A szaporodás módja kettéosztódás (binary fission), ami NEM mitózis, hiszen sem cenriólumok, sem kinetokór mikrotubulusok sincsenek (a kromoszómákról, illetve kromatidákról már nem is beszélve).
Az energianyeréshez a prokarióták már felhasználják a H+ elektrokémiai potenciál-különbséget, tehát működtetnek ATP-szintázt a plazmamembránban. A protonok felhalmozási helye a Gram-negatív baktériumokban a periplazmikus tér, azaz a plazmamembrán és a külső membrán közötti tér. A Gram-pozitív baktériumokban a sejtmembrán és a sejtfal közötti térben akkumulálódnak a protonok, és hasonló a helyzet az Archaeákban is. A lényeg azonban az, hogy az energianyerés alapja a proton elektrokémiai különbség létrehozása és ennek energiájával ATP szintézise.
Mozgásuk flagellum(ok) segítségével történik, amely(ek) a plazmamembránból kiinduló képlet(ek), tehát nem sejtalkotók, mint az Eukariótáknál (ott ugyanis a flagellumot kívülről körülveszi a plazmamembrán, tehát ott a flagellum valódi sejtalkotó.
Szignalizációs rendszereik két-komponensű rendszerek, amelyek szignál-receptor kinázból és válasz-regulátorból állnak. A szignált felfogó receptor egyszersmind egy kináz, amely először saját magát foszforilálja (autofoszforiláció) egy hisztidin (His) aminosavon, majd ez a foszfátcsoport átkerül a válasz-regulátor egyik aszparaginsav (Asp) aminosav-oldalláncára.
Táplálkozásukra jellemző kémiai molekulák környezetből történő felvétele, de nem jellemző egy másik egyed bekebelezése, azaz a fagocitózis.
Felmerül a kérdés, hogy amennyiben a baktériumok jelen vannak a Földön az első sejt kialakulása óta (kb. 3,8 milliárd éve), vajon ez a mérhetetlenül hosszú evolúciós idő alatt miért nem változtak jelentősen sejtfelépítésüket tekintve. Ha megfigyeljük, a fejlettebb eukariótákkal összehasonlítva a baktériumokra (prokariótákra) alapvetően két jelző alkalmazható: kis testméret és a komplexitás hiánya. Annak a ténynek, hogy ezek a sajátosságok nem változtak csaknem 4 milliárd évnyi fejlődés során (vagy ha úgy tetszik ezek a jellegek konzerválódtak az öszzes prokariótánál), valami oka kell, hogy legyen. Ennek vizsgálatára tekintsük át a baktériumok testfelépítését és táplálkozásmódjukat. A baktériumok belső szerveződése strukturálatlan, a sejt belső térfogatát a homogén citoplazma tölti ki, benne legfeljebb a cirkuláris DNS kópiáit, plazmidokat és a citoszkeleton előfutárait találjuk. A sejt a tápanyagokat a sejtfelszínen veszi fel, az életfolyamatok viszont a teljes belső állományban zajlanak, tehát nem mindegy, hogy milyen térfogatú belső állományt kell a sejtfelszínnek (sejtmembrán) tápanyaggal ellátnia. Minél nagyobb a sejtfelszín területe (A = area) a sejt térfogatához (V = volume) viszonyítva (magas A/V arány), annál hatékonyabb a sejt működése, ugyanis időegység alatt kevesebb tápanyagot kell felvennie azonos térfogatú citoplazma „kiszolgálásához”. Ha a sejt növekszik, a felszín kevésbé gyorsan nő, mint a térfogat, ami a kedvező A/V arányt kedvezőtlen irányba tolja. Gömbalakú baktériumok (coccusok) esetén a felület: A = 4 r2 π, a térfogat: V = (4 r3 π)/3, az A/V arány tehát 3/r. Tegyük fel, hogy egy coccus sugara 1 μm, ekkor a felszín/térfogat aránya 3. Ha a sejt növekedésbe kezd, és a sugara megduplázódik, akkor a sejt felszín/térfogat aránya már csak 1,5 lesz, azaz azonos térfogatot már csak feleakkora felszín tud kiszolgálni. További növekedéskor a felszín/térfogat arány még kedvezőtlenebbé válik, ami a sejtet arra készteti, hogy valamit tegyen a kedvezőtlen felszín/térfogat arány helyreállítására. A legegyszerűbb és leghatékonyabb megoldás, ha a sejt megfeleződik, és utána minden kezdődik elölről. Mindez egyszerűen és szemléletesen magyarázza, hogy a sejtméret növekedése miért előnytelen egy baktériumsejt számára (31. ábra).

31. ábra
A prokarióták gyors osztódási rátája tehát egyrészt következik a táplálkozási szokásaikból. A baktériumsejt osztódása nem mitózis, hiszen nincsenek valódi kromoszómák, sem pedig kinetokór mikrotubulusok. Hivatalosan kettészakadásnak (binary fission) nevezik. Az eukariótákhoz hasonlóan, viszont, a kettészakadás előtt a bakteriális genom is megkettőződik, és ez a másik kulcsa a baktériumok extrém gyors osztódásának (32. ábra).
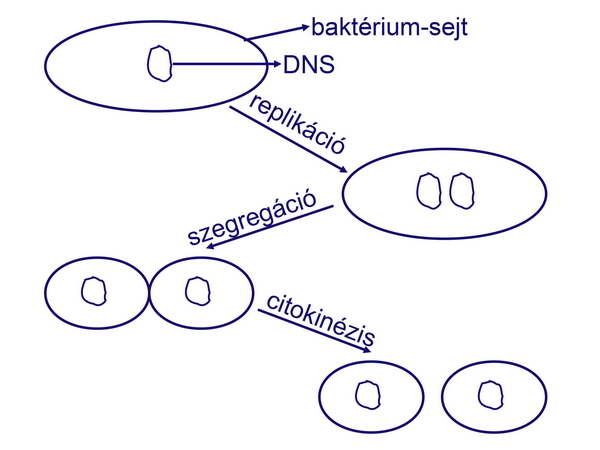
32. ábra
Vida Gábor Professzor (ELTE Genetikai Tanszék) és kutatócsoportja végezte el azt az érdekes kísérletet, amely megmagyarázza a baktériumok gyors osztódását. E.coli baktériumokat transzfektáltak különböző méretű plazmidokkal, amelyek hordoztak egy antibiotikum-rezisztencia gént. Ha a baktériumokat olyan táptalajra szélesztették, amelyben jelen volt az antibiotikum, az a törzs osztódott a leggyorsabban, amelyben a plazmid mérete a legkisebb volt. Ha azonban a törzseket olyan táptalajra szélesztették, amelyben nem volt antibiotikum, valamennyi törzs azonos sebességgel osztódott. Kiderült, hogy a baktériumok az antibiotikum távollétében a „felesleges” géneket hordozó plazmidoktól megszabadultak a minél kisebb genom-méret, azaz minél gyorsabb osztódás érdekében. A kolónia „fitness”-ét (azaz szaporodási sikerét) ugyanis annak osztódási sebessége szabja meg: ezt az elképzelést azóta „use it or lose it” hipotézisként ismeri a tudománytörténet.