Tartalom
Az első sejt megjelenése a Földön kb. 3,5-3,8 milliárd évvel ezelőtt következhetett be. Ez olyan hatalmas időszak, hogy hétköznapi eszünkkel nem sokat tudunk mit kezdeni vele. Képzeljük el azonban, hogy egy olyan rakétában utazunk, amely 1000 évet tesz meg másodpercenként. Ebben az időrakétában 10 másodperc utazás után már látnánk az utolsó jégkorszakot és pl. a Balaton keletkezését, de 18 órát kellene utazni ahhoz, hogy tanúi legyünk a Dinoszauruszok kihalásának, 4 és fél napot ahhoz, hogy lássuk az első többsejtűeket, 24 napot ahhoz, hogy tanúi legyünk az első eukarióta sejt megjelenésének, és 44 nap utazás után találkozhatnánk LUCA-val.

1. ábra
Az élet korai megjelenésére geológiai bizonyítékok és prekambriumi fosszíliák utalnak többek között olyan fosszilis sztromatolitok, amelyek korát 3,5 milliárd évre becsülik (2. ábra).
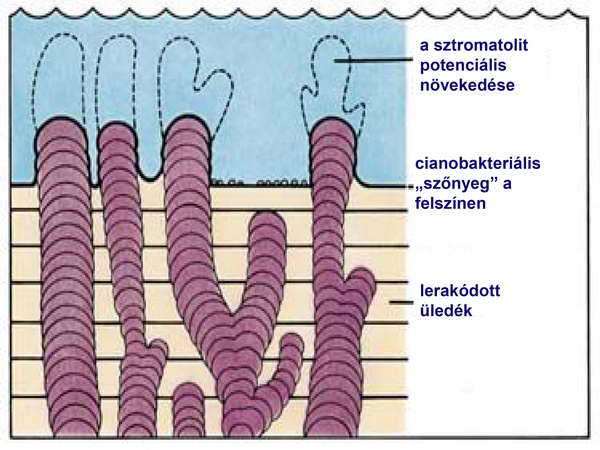
2. ábra
Léteznek récens sztromatolitok is, többek között Ausztrália partjai mentén (3. ábra).

3. ábra
A sztromatolitokat az „A sejtek információhordozó rendszereinek evolúciója” című fejezetben taglaljuk. Az uralkodó elképzelés szerint ebben a korai időszakban a Föld ősi atmoszférája redukáló volt, amelyben dominált az ammónia (NH3), metán (CH4) és a szén-dioxid (CO2), hiányzott viszont belőle az oxigén (O2). Ezt az elképzelést az 1910-es években alkotta meg egymástól függetlenül egy orosz (Oparin) és egy brit (Haldane) tudós. Mindketten a Naprendszer legnagyobb, és egyik légkörrel rendelkező bolygója, a Jupiter légkörében levő gázok összetételét vizsgálták olyan távcsövekkel, amelyekkel színképelemzési módszerekkel a gázok elemzését és azonosítását el lehetett végezni. (4. ábra).

4. ábra
Mivel ott a légkörben főleg az említett gázokat találták, azt posztulálták, hogy az ősi Föld légköre is hasonló összetételű volt. Mindez tápot adott további hipotéziseknek, amelyek egyike Miller híres kísérletéhez vezetett, amit az akkor már Nobel-díjas Urey (a deutérium felfedezője) laboratóriumában végzett. A kísérletben egy lombikban elegyítette ezeket a gázokat, majd elektródákkal elektromos kisüléseket indukált, és a lombik alján levő folyadékban szerves anyagok (aminosavak) jelenlétét mutatta ki. (5. ábra)
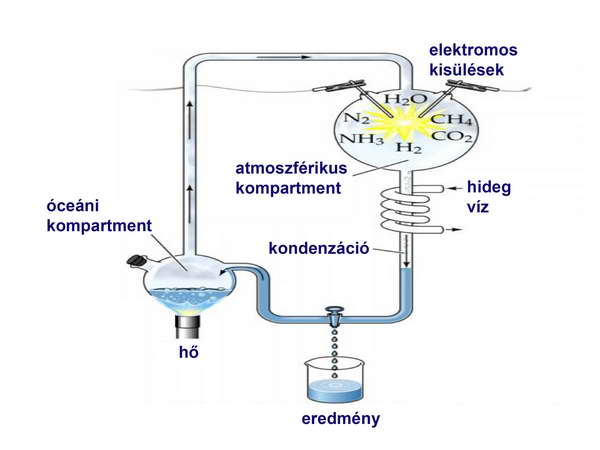
5. ábra
Ez igen fontos megfigyelés, hiszen igazolni látszott, hogy a fehérjéket felépítő aminosavak és esetleg az élő anyag további építőkövei (pl. nukleotidok) létrejöhettek abiogén úton a Föld ősi reduktív atmoszférájában. Az akkoriban igen híres és „korszakalkotó”-nak számító kísérletet azonban másoknak nem sikerült megismételni, és ez jelentette az első komoly kihívást az ún „Őslégkör-teóriá”-val kapcsolatban.
Értelemszerű, hogy az első elméletek a korai sejtek energianyerésével kapcsolatban abból indultak ki, hogy mivel az őslégkör redukáló volt, nem volt oxigén a sejtek közelében, ezért azok kizárólag anaerob módon, fermentációval nyerték az energiát, azaz szubsztrát-szintű foszforilációval termelték az ATP-t (6. ábra), és ez a típusú energianyerés volt az ATP-gyártás domináns módja egészen addig, amíg az első cianobaktériumok meg nem jelentek és az oxigént termelő fotoszintetikus ATP-szintézist meg nem valósították.
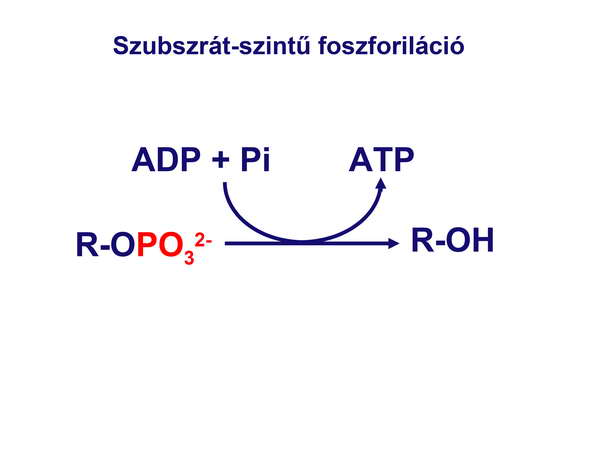
6. ábra
Mivel az Őslégkör-teória szerint a cianobaktériumok előtt nem volt oxigén a Földön, kizárt volt az a lehetőség, hogy már az első sejtek oxidatív energianyerésre lettek volna képesek. Tehát a korai elképzelések szerint először fermentatív („szubsztrát-szintű”) volt az ATP-szintézis, majd a cianobaktériumok hozták az első kemiozmotikus mechanizmust, és ezt követően a melléktermékként keletkező oxigén feldúsulásával fokozatosan álltak át a sejtek mitokondriumai az oxidatív foszforilációra.
Ez a teória igen logikusnak és elfogadhatónak tűnt. Nagy hibája azonban, hogy nem adott elfogadható magyarázatot a kemiozmózis hirtelen, „ugrásszerű” megjelenésére a cianobaktériumoknál. A kemiozmotikus energianyerés annyira alapvetően különbözik a szubsztrát-szintű foszforilációtól, hogy nehezen elképzelhető, hogy mindössze egyetlen progresszív evolúciós mozzanat eredménye. A kemiozmotikus energinyerés alapja, hogy egy membránnal elkerített térrészben ionok halmozódnak fel, azaz elektrokémiai potenciál-különbség alakul ki a membrán két oldalán. Ez a grádiens magas szabadenergia-tartalommal rendelkezik (lásd „A biológiai energianyerés lényege” fejezetet), amelyet egy fordított üzemmódban működő ATPáz (azaz ATP-szintáz) a biológiai energiahordozó, az ATP szintézisére tud fordítani (7. ábra).
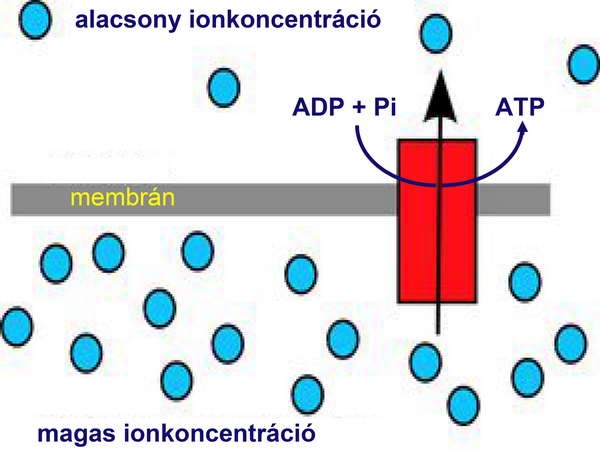
7. ábra
Még kevésbé magyarázható meg az a tény, hogy ugyanaz a mechanizmus (kemiozmotikus energianyerés) hogyan kerülhetett bele a mitokondriumok ősébe, amelyek a cianobaktériumoknál primitívebb prokarióták voltak, ráadásul úgy, hogy a mitokondriumok csak és kizárólag oxigén jelenlétében működnek, viszont a Föld korai körülményei között a teória értelmében nem volt oxigén? Mindez csak úgy képzelhető el, hogy ez az energianyerési mód már jelen kellett, hogy legyen abban a sejtben, amelyből mind az ősi mitokondriumok, mind a későbbi cianobaktériumok származtak. Mindezek igen komoly érvek, amelyekkel szemben az Őslégkör teória nem tudott felvonultatni meggyőző ellenérveket.
Ezen túlmenően még komolyabb kihívások is érték a teóriát a modern molekuláris módszerek részéről. Kiderült ugyanis, hogy mind a récens Eubaktériumok, mind az Archaebaktériumok hordoznak olyan géneket, amelyek fehérjetermékei antioxidáns enzimek (szuperoxid dizmutáz, kataláz). A kataláz ribbon-diagrammját a 8. ábrán mutatjuk be (8. ábra).
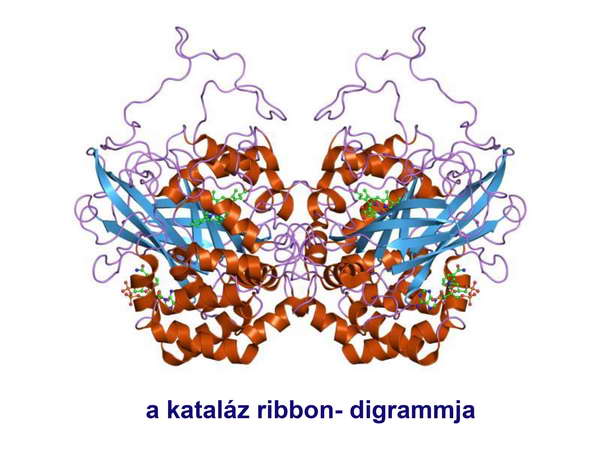
8. ábra
Mindez jelezte azt, hogy a korai prokarióták már rendelkeztek olyan enzimatikus mechanizmusokkal, amelyek felvértezték őket az oxigén szabadgyökök ellen és lehetővé tették számukra e veszélyes szabadgyökök detoxifikálását. Az oxigén speciális molekuláris szerkezetéről és szabadgyök-képző hajlamával bővebben az „A sejtek információhordozó rendszereinek evolúciója” című fejezetben foglalkozunk. Még nagyobb tudományos szenzáció volt, amikor 1996-ben Castresana és Sarastre publikálta azt a megfigyelésüket, hogy a terminális oxidáció végső lépését katalizáló citokróm oxidáz enzim génjét kimutatták mind récens Eubaktériumokból, mind Archaebaktériumokból. Ez ugyanis feltette a koronát az őslégkör-teória bírálatára, hiszen ez azt jelentette, hogy mindkét prokarióta csoport nemcsak védekezni tudott az oxigén káros hatásai ellen, hanem hasznosítani is tudta azt, ráadásul pontosan ugyanúgy, ahogy azt a mai mitokondriumaink teszik, azaz aerob környezetben.
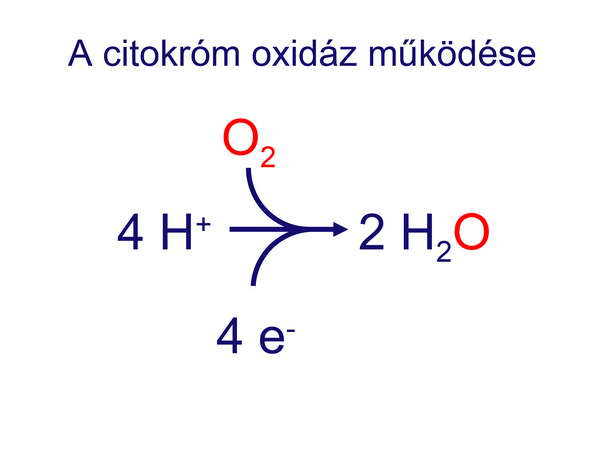
9. ábra
A citokróm-oxidáz által katalizált rekció részleteit a 10. ábrán mutatjuk be.
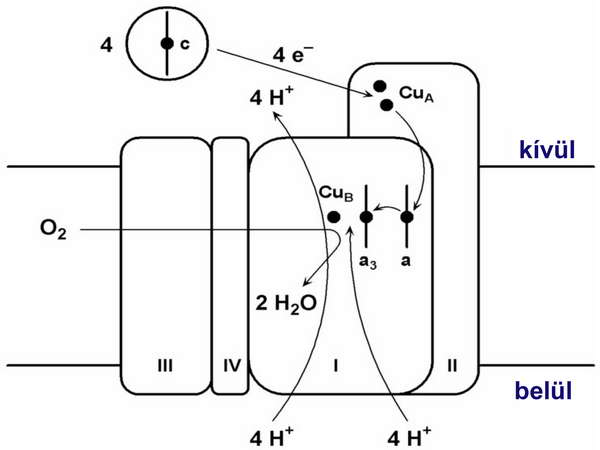
10. ábra
Mindezeken túlmenően, ez a felfedezés azt is bizonyossá tette, hogy az evolúció során korábban alakult ki az oxidatív energianyerés, mint a fotoszintézis, ami egy újabb ponton mondott ellent az Őslégkör hipotézisnek.
Akkor viszont felmerül a kérdés, hogy a Föld ősi körülményei között hogyan keletkezhetett az oxigén? Az Őslégkör teória szerint az oxigén csak a cianobaktériumok fotoszintetikus aktivitásával jelent meg a Földön, de ennek az újabb molekuláris bizonyítékok ellentmondanak. A cianobaktériumok előtt viszont nem élt olyan prokarióta, amely oxigént tudott volna termelni, hiszen sem az Archaebaktériumok, sem a korábbi Eubaktériumok nem képesek oxigén-termelő fotoszintézisre. A legelfogadhatóbb magyarázat szerint az oxigén abiogén módon, a Földre akadálytalanul érkező nagy-energiájú UV-sugarak közreműködésével keletkezett, amelyek bontották az ős-tenger vízmolekuláit H2-re és O2-re (fotolízis). Ez egy olyan láncreakció, amelyben az oxigén négy lépésben keletkezik. A vízmolekula minden lépésben veszít egy-egy elektront,
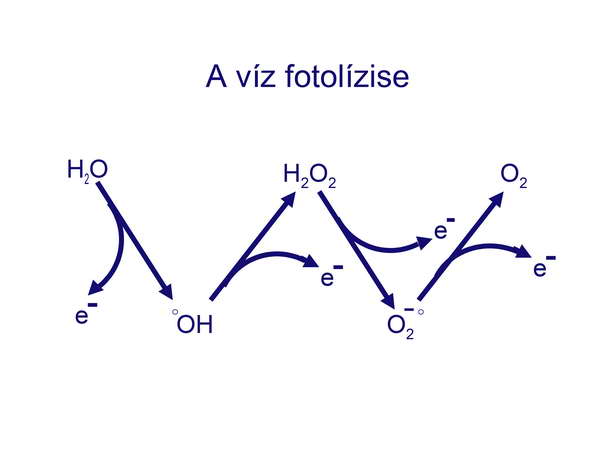
11. ábra
amelyek a négy protonnal két hidrogén molekulává rekombinálódnak. A hidrogén a legkönnyebb gáz, tehát azonnal elillant a légkörből, az oxigén viszont itt maradt és bár csekély, de mérhető mennyiségben bekerült az ős-óceán vizébe oldott formában. Becslések szerint az oxigén akkori koncentrációja 0,1% lehetett a légkörben, amely a cianobaktériumok tevékenysége révén érte el a 2%-ot, de még az első többsejtűek megjelenésekor (kb. 800 millió évvel ezelőtt) is mindössze 5 % volt.

12. ábra
(Megjegyezzük, hogy a „kambriumi robbanás” során a légköri oxigén koncentrációja elérhette a 35%-ot is, egyes elképzelések szerint ez vezetett a „gigászok” megjelenéséhez a karbon-perm-jura időszakokban, azaz mintegy 350-től 100 millió évvel ezelőttig. Ezek között a dinoszauruszok mellett pl. 70 cm fesztávolságú szitakötők, 1 m hosszúságú skorpiók, valamint 5 m-es kételtűek is éltek).

13. ábra
Az újabb elképzelések szerint a víz fotolízise mindaddig tartott, amíg a cianobaktériumok által termelt oxigénből nem keletkezett elég ózon, amely megakadályozta az UV-sugarak Föld-felszínre érkezését, és ezzel megmentette a Földet attól, hogy olyan sorsa jusson, mint pl. a Mars. A legújabb műhold-felvételek egyértelműen bizonyítják, hogy a Marson is volt víz (sőt állóvíz is), viszont a fotolízis hatására az összes víz elbomlott. További bizonyíték erre az a tény, hogy a víz fotolíziséből keletkező oxigén a Marson oxidálta a felszíni kőzeteket, többek között a vasat, aminek eredményeként a vörös ferri-kőzetek dominálnak a bolygón, indokolva annak csillagászati elnevezését (vörös bolygó).

14. ábra
Tehát, a Földön a cianobaktériumok fotoszintézise mentette meg az ős-óceán vizét a fotolízistől!
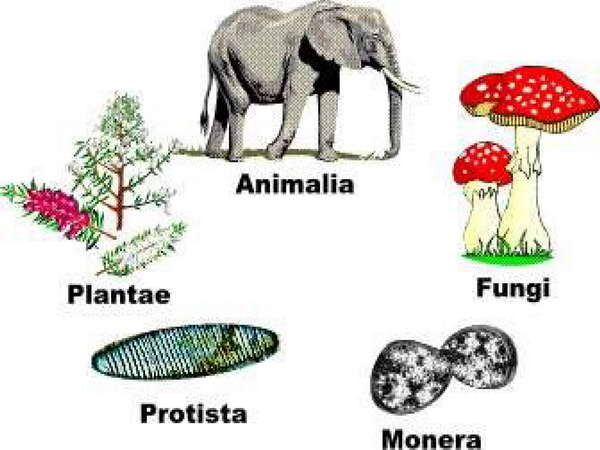
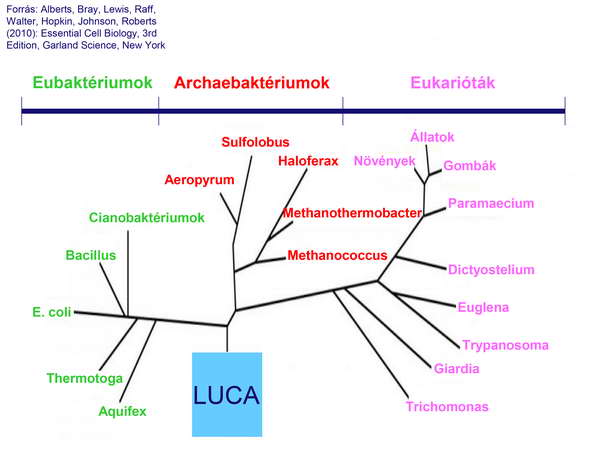
A jelenlegi hallatlanul formagazdag élővilág láttán paradoxonnak tűnik első közös sejtről beszélni. A fajok fokozatos kialakulására vonatkozó darwini koncepció azonban már felvetette, hogy az élet létrejötte a Földön csak egyszer következett be, és minden mai, valamint fosszilis élőlény ennek a hipotetikus ősi élőlénynek (őssejtnek) a leszármazottja. A valódi áttörést azonban Woese 1977-ben publikált korszakalkotó felfedezései (az Archaebaktériumok (Archaeák) „újrafelfedezése” és főleg az „Élet fája” modell elkészítése) jelentették. Ez a kísérleti bizonyítékokon alapuló (a riboszómális 16S sRNS génjének szekvencia-analízise alapján felállított) törzsfa ugyanis végérvényesen szakít a korábbi „5 ország” modellel (Állatok, Növények, Gombák, Eukarióta egysejtűek (Protiszták) és Prokarióták),
15. ábra
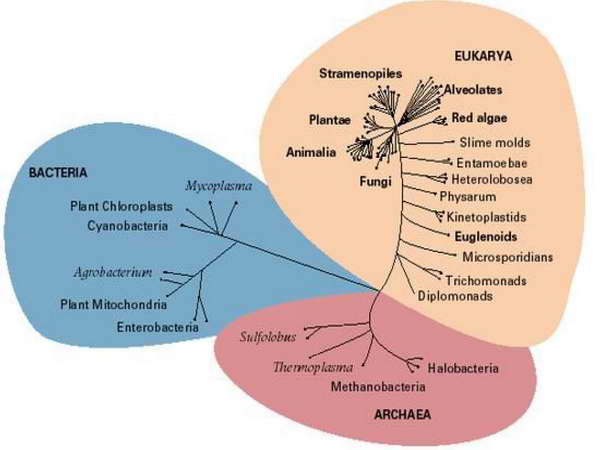
és az élővilágot 3 fő doménba sorolja (Eubaktériumok, Archaebaktériumok és Eukarióták.
16. ábra
17. ábra
Ez a forradalmi modell azt is egyértelművé teszi, hogy a Prokarióták korábban váltak le a fejlődési vonalról, mint az Eukarióták, aminek az a hallatlanul meglepő és érdekes következménye, hogy az Eukarióták közelebbi rokonságban állnak az Archaebaktériumokkal, mint az Eubaktériumokkal. Harmadik fontos következtetése a modellnek, hogy a fejlődési vonalon visszafelé haladva fordított irányban tudjuk követni az evolúció folyamatát, és emiatt az Eubaktériumok és az Archaebaktériumok közös ősének a léte a hipotézisből a realitás talajára került. Ezt a közös őssejtet az angol irodalomban Last Universal Ancestornak (Utolsó Egyetemleges Ősnek) vagy Last Universal Common Ancestornak (Utolsó Egyetemleges Közös Ősnek) nevezik, és ma már lehetőségünk van felvázolni ennek a csaknem 4 milliárd évvel ezelőtt élt sejtnek a legfontosabb tulajdonságait.
A közös ősök tanulmányozása a darwini evolúciós elveken alapul: számba kell venni a leszármazottakban fellelhető közös tulajdonságokat, és ezek alapján nagy valószínűséggel megjósolhatjuk a közös ős jellegzetességeit is. Ezen deduktív következtetések alapján tehát LUCA-ról nagy valószínűséggel az alábbi jellemvonások tételezhetők fel:
Egysejtű prokarióta szervezet volt, mérete kb. azonos a mai prokariótákéval (kb. 1-2 μm).
A sejtet legkívül sejtfal határolta.
Mutatta a sejtes szerveződés minden fontos kritériumát: külső határolóhártya (membrán), benne a sejt belső állománya (citoplazma), ebben cirkulárisan rendeződött örökítő anyag.
A citoplazma ionösszetételére jellemző volt a magas fehérjetartalom, az alacsony Na+ és a magas K+ koncentráció.

18. ábra
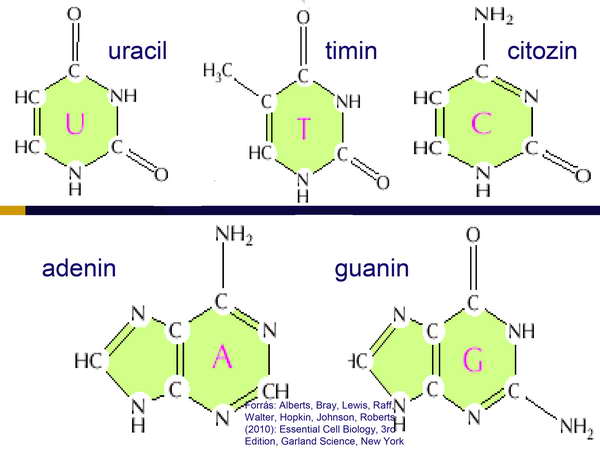
A sejt genetikai állománya DNS alapú volt, melyet adenin, timin, guanin és citozin bázisok alkotta nukleotidok építettek fel.
19. ábra
A kétszálú DNS replikációjáról templát-fűggő DNS-polimeráz gondoskodott
A DNS integritásának fenntartásáról DNS topo-izomerázok, ligázok és javító (repair) enzimek gondoskodtak
A DNS mellett rendelkezett mindhárom típusú RNS molekulával (mRNS. tRNS, rRNS), melyekben a timin helyett uracil bázis fordult elő (19. ábra).
A hírvivő (messenger) RNS (mRNS) molekulák létrejöttét a DNS-függő RNS-polimeráz katalizálta a DNS nukleotid-sorrendje alapján
A sejt citoplazmájának alapanyagát főleg fehérjemolekulák alkották, amelyek 20 aminosavból felépülő 3-dimenziós polipeptid-szerkezetet hoztak létre. A fehérjékben csak az aminosavak L-módosulatai fordultak elő.
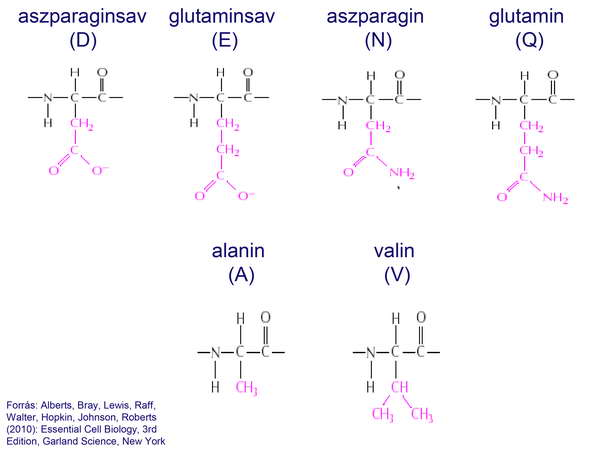
20 ábra
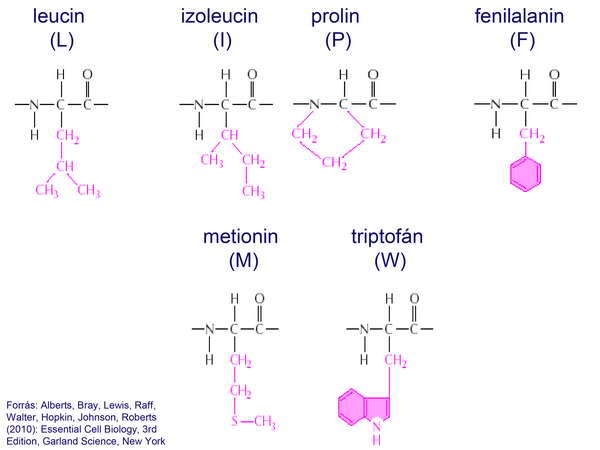
21. ábra
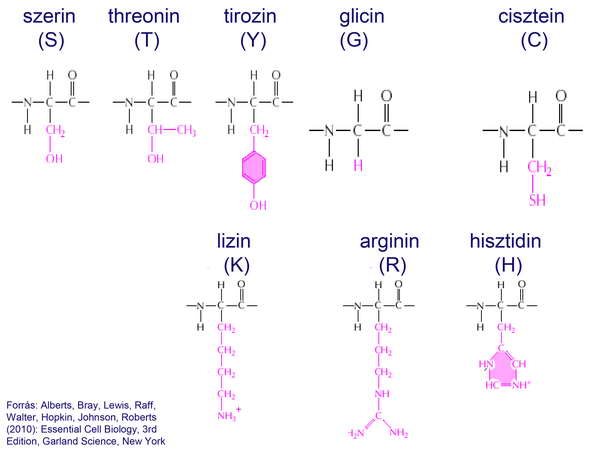
22. ábra
A fehérjék között már jelen voltak a DNS-bioszintézis, RNS-bioszintézis és a fehérje-bioszintézis enzimei.
A DNS-ben rögzült információ két lépésben (transzkripció és transzláció) fejeződött ki és egy új fehérje megjelenését eredményezte a sejtben.
A fehérjék felépítéséhez ugyanazt a típusú genetikai kódot használta, ami az összes mai élőlényben is univerzálisan használatos.
A fehérjék 70S szedimentációs állandójú riboszómákon készültek el a citoplazmában, amelyek egy nagyobb (50S) és egy kisebb (30S) alegységből álltak.
Az anyagcsere és az energiagyártás alapját szerves molekulák képezték, amelyeket membránján keresztül vett fel és hasznosított.
Energiaigényes folyamatait ATP-vel fedezte és képes volt az ATP előállítására.
Energianyerésének hátterében a redoxi folyamatok álltak, azaz már képes volt az elektronátmeneti energiát (elektromos feszültség-különbség) átalakítani az ATP kémiai kötéseiben tárolt energiává. Mind az elektron-donort, mind az elektron-akceptort illetően számos melekula vagy ion valószínűsíthető. Utóbbiak között már szerepel az oxigén is.
Az ATP-szintézist kemiozmotikus mechanizmussal végezte: rendelkezett olyan H+-ATPázzal, amely a lokális proton elektrokémiai potenciál-grádienst használta fel, ez fedezte a megfordítva működő H+-ATPáz (ATP szintáz) számára az ADP ATP-vé történő foszforilációjának energiaigényét. (23. ábra)
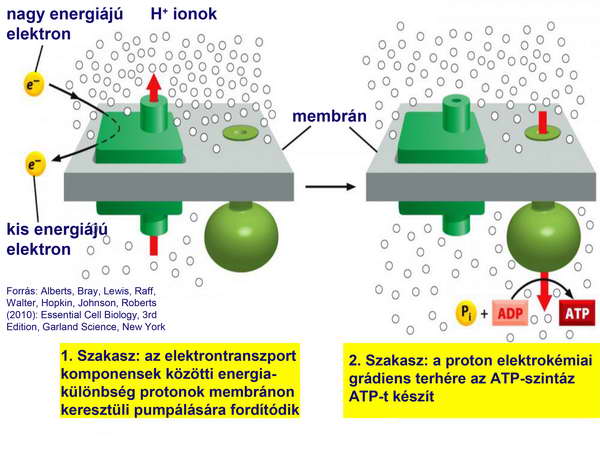
23. ábra
Az ATP szintézisét megvalósító enzim-komplex, az ATP-szintáz, tulajonképpen egy fordított irányban működő ATPázról van szó. Ez az ATPáz az ún. F-típusú (vagy F0-F1) ATPázok közé tartozik (a mitokondriumok belső membránjában, a kloroplasztiszok tilakoidmembránjában és a baktériumok membránjában található H+-ATPázok összefoglaló neve), amelyek több alegységből állnak. Ezek funkcionálisan két alapvető tevékenységet végző részre oszthatók: egy H+-csatornára és egy ATPázra, ami a koncentráció-viszonyok miatt nem az ATP-hidrolízis (bontás), hanem az ATP-szintézis irányában működik. A H+ elektrokémiai potenciál-különbség hajtóereje révén a megnyíló H+-csatornán keresztül a protonok beáramlanak a mitokondrium mátrixába, és ezzel egyidejűleg a felszabaduló energia terhére az ATP-szintetáz ADP-ből és szervetlen foszfátcsoportból ATP-t szintetizál, tehát a protonok elektrokémiai potenciál-különbsége nagyenergiájú foszfátkötések létesítésére fordítódik (24. ábra). E kemiozmózisnak nevezett mechanizmust Peter Mitchell fejtette meg, aki ezért a korszakalkotó felismeréséért Nobel-díjat kapott.
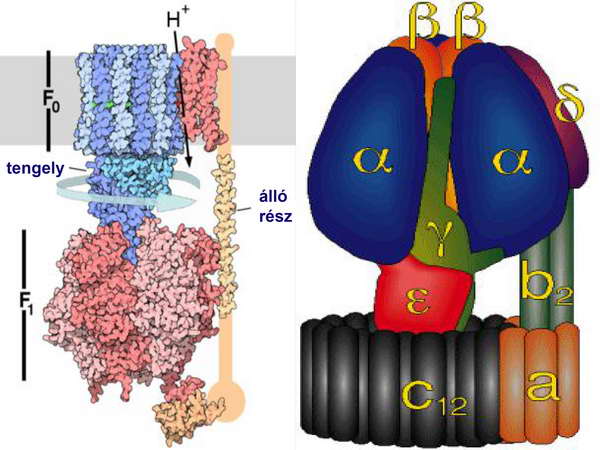
24. ábra
Tudott védekezni az oxigén szabadgyökök ellen, tehát már termelte is azokat.
Mindezek a tulajdonságok szinte minden élő sejt alaptulajdonságai, amelyekkel a deduktív logika szerint már LUCA is rendelkezett. Vannak azonban ismeretlen részletek, „fehér foltok”, amelyek kiszínezése még várat magára. Több olyan részlet van, ugyanis, amire nincs egyértelmű válasz. Ezek közé tartoznak azok a sajátosságok, amelyekben az Eubaktériumok és az Archaebaktériumok eltérnek egymástól, gyakran elég jelentősen. Ezek közül a legfontosabbak a következők:
Miből állt LUCA sejtfala? Az Eubaktériumoknak peptidoglukán, az Archaebaktériumoknak fehérje-alapú sejtfaluk van (pszeudo-murein), ezért a közös ős sejtfala valami olyasféle struktúra kellett, hogy legyen, amelyből mind két típus kialakulhatott.
Miből állt LUCA sejtmembránja? Ez még nehezebb kérdés, hiszen az Eubaktériumok és az Archaebaktériumok membránja gyakorlatilag semmiben sem hasonlít egymásra. Az Eubaktériumoknál a membránt foszfolipidek építik fel, melyben a glicerin alapmolekulához két apoláros zsírsav és egy poláros fejcsoport kapcsolódik észter-kötésekkel. Az Archaebaktériumok membránjában a glicerinhez nem zsírsavak, hanem poli-izoprén láncok kapcsolódnak, ráadásul éter-kötésekkel. Eltérő a lipidek szteroizomériája is. Egyszóval, a két utódsejtben található két gyökeresen eltérő membrán-összetétel ismeretében igen nehéz rekonstruálni a közös ős-sejt sejtmembránjának összetételét.
Ennek az igen komoly dilemmának a feloldására fejlesztette ki teóriáját Martin és Russel, akik az élet megjelenését a vas-kén világban valószínűsítik. Elképzeléseik szerint az első sejteknek nem is volt a mai értelemben vett membránjuk, hanem helyette szervetlen anyagok (vas-kén ásványok) alkothatták az első élő sejt membránját. A teória épít Günther Wächtershäuser korábbi hipotézisére, aki szerint az élő sejtek első katalitikus rekciói még a fehérje-előtti időszakban is elképzelhetők voltak, katalizátorként a vas-kén ásványok szerepeltek. Ebben a világban még nem volt membrán, hanem csak katalitikus folyamatok, amelyek a tengerek mélyén elhelyezkedő hidrotermális kürtőkben, az ún. fekete füstölőkben (black smokers), pontosabban azok külső felületén mentek végbe. A fekete füstölők speciális mélytengeri geológiai képződmények, amelyekben speciális körülmények (magas nyomás és magas hőmérséklet, akár 100 Co) uralkodnak. Több ezer méteres mélységben a tengerből a tengerfenék kőzetébe beszivárgó vízet a magmából érkező hő felmelegíti, de a nagy nyomás miatt a víz forráspontja is emelkedik („superheating”), tehát az folyékony halmazállapotú marad 100 Co közelében is. A tengerfenék felszíne alatt összegyűlt víz a nagy nyomás következtében olyan helyeken összpontosul, ahol a kőzet lazább, és ott áttöri a tengerágyat és tengervízbe jut. Itt viszont már alacsonyabb a hőmérséklet, ezért a vízben oldott gázok kilépnek és gáz formában haladnak a felszín felé (25. ábra).

25. ábra
Ezek a gázok főleg szén-dioxidból és kénhidrogénből állnak. Az igazi meglepetést az váltotta ki, amikor kiderült, hogy ezekben a récens hidrotermális kürtőkben a szó szoros értelmében pezseg az élet a meghökkentő körülmények (magas nyomás, magas hőmérséklet, teljes sötétség) ellenére! Az elképzelés értelmében vas-kén ásványok működtek katalitikus centrumként és felületükön katalizálták a szén megkötését, azaz egyszerű szerves molekulák abiogén létrejöttét a szervetlen gázokból (CO2, CO, HCN és H2S). Ezek a primitív szerves molekulák idővel a metallo-peptidek megjelenéséhez vezettek, amik a mai citokrómok, SOD és más fontos enzimek előfutárai voltak.
LUCA létezése ma már kilépett a tudományos-fantasztikum világából és mintegy 4 milliárd évvel ezelőtt valóságosan élt realitássá vált. Ez hallatlanul fontos az élet kialakulásának megértése és rekonstruálása szempontjából. A molekuláris módszereknek köszönhetően egyre többet tudunk felépítéséről és életmódjáról, de vannak még megválaszolandó kérdések, amikre a tudomány remélhetőleg már a közeljövőben választ ad.
Milyen megfigyelések alapján jött létre az „Őslégkör hipotézis”?
Ha egy „időrakétában” 1000 év/s sebességgel utazunk visszafelé az időben, mennyit kellene utaznunk ahhoz, hogy találkozzunk LUCA-val?
Mi a kemiozmotikus energianyerés lényege?
Mi a szubsztrát-szintű foszforiláció lényege?
Milyen molekuláris bizonyítékok szólnak amellett, hogy az ősi Földön már kellett, hogy legyen oxigén?
Milyen reakciót katalizál a citokróm oxidáz?
Hogyan keletkezik az oxigén a víz fotolízise során?
Milyen sorsa jutott a Mars a Földdel ellentétben és miért?
Miért volt forradalmi Woese „Az élet fája” modellje?
Mi alapján állította fel Woese az „Az Élet Fája” modellt?
A mai sejtekben hol találunk F0-F1 ATP-ázokat?
Miért valószínű, hogy LUCA-nak volt sejtfala?
Milyen bizonytalanság uralkodik LUCA sejtmembránjával kapcsolatban?
Mik azok a „fekete füstölők” és hogyan járulhattak hozzá az élet keletkezéséhez?
Mely kutatók nevéhez fűződik a „vas-kén világ” hipotézis?