Tartalom
A prokarióta és az eukarióta sejtek közötti különbség a méret és a komplexitás vonatkozásában annyira szembetűnő, hogy nehéz elképzelni, hogy egyáltalán van valami közük egymáshoz. Az eukarióta sejtek óriások a prokariótákhoz viszonyítva, és bennük számos sejtorganellum, citoszkeletális struktúrák és belső membránrendszer is található a citoszol és a DNS mellett. Kialakulásuk mintegy 2 milliárd évvel ezelőtt következett be, emiatt ennek történései nehezen rekonstruálhatók, és így a kutatók kísérleti bizonyítékok hiányában elméleteket állítanak fel a legvalószínűbb forgatókönyvekről. A legnagyobb érdeklődés és vita a saját genommal rendelkező sejtorganellumok (mitokondriumok és kloroplasztiszok) eredete körül bontakozott ki. Ez a kérdés régóta foglalkoztatja a kutatókat. Először a kloroplasztiszokat vették szemügyre, hiszen azok jóval nagyobbak (5-20 μm), mint a mitokondriumok (1-3 μm). Már az 1800-as évek végén (!) kiderült, hogy a kloroplasztiszok saját életciklust követnek, ezen belül osztódnak, majd arra is fény derült, hogy osztódásuk nem az eukarióta mitózishoz, hanem a cianobaktériumok osztódásához (húzófonalak nélküli kettéhasadás) hasonló. Egy jó évszázaddal később, az 1920-as években hasonló megfigyeléseket tettek a mitokondriumokkal is. Később az elektronmikroszkóp fontos strukturális hasonlóságokra világított rá egyrészt a kloroplasztiszok és a cianobaktériumok, másrészt a mitokondriumok és a baktériumok között. Az 1970-es években pedig kiderült, hogy mind a mitokondriumok, mind a kloroplasztiszok rendelkeznek egyrészt önálló DNS-sel, másrészt saját riboszómákkal is, ráadásul olyanokkal, amelyeket korábban csak a prokariótákban találtak meg.
Ezek alapján alakult ki az az elképzelés, hogy az eukarióta sejtek úgy alakultak ki, hogy korábbi primitívebb sejttípusok bekebeleztek prokarióta szervezeteket, és ezt követően tartósan együtt maradtak. Mivel tartós együttélés és kölcsönös előnyökkel járó együttműködés bontakozott ki a bekebelezett (szimbiont) és a bekebelező (gazda) sejtek között, ezt a fajta kooperációt „endoszimbiózis”-nak nevezték el. A folyamatot leíró elképzelés zászlóvivője és egyik legfőbb hirdetője Lynn Margulis (egy hölgy), akinek 1981-ben megjelent „Symbiosis in Cell Evolution” című könyve összegzi az azóta „endoszimbionta elmélet”-nek elnevezett bizonyítékait és felvázolja annak valószínű eseménysorát. Maga az endoszimbiózis ténye és a két sejtalkotó (mitokondrium és kloroplasztisz) endoszimbionta (azaz a baketriális őstől származó) eredete kétségbevonhatatlanná vált, és a mai sejtbiológiai gondolkodásunk alapját képezi. Ennek bizonyítékai:
mind a mitokondriumok, mind a kloroplasztiszok rendelkeznek saját DNS-sel, amely alakjában hasonlít a bakteriális DNS-re,
mind a mitokondriumok, mind a kloroplasztiszok 70S szedimentációs állandójú riboszómákat tartalmaznak, azaz olyanokat, mint amik prokarióta sejtekben találhatók,
a sejtmagi DNS-ben szép számmal találhatók mitokondrium- és kloroplasztisz-eredetű gének, amelyek organelláris eredetűek és a legvalószínűbb forgatókönyv szerint horizontális gén-transzferrel kerültek át a nukleáris genomba,
mindkét sejtorganellumot legalább két teljes membrán határolja, melyek eltérő összetételűek és eltérő funkciójúak,
mindkét sejtorganellum képes önálló osztódásra, amely emlékeztet a baktériumok kettéosztódására,
mind a kloroplasztiszban, mind a mitokondriumban találunk olyan fehérjéket és elektrontranszport-lánc komponenseket (pl. citokrómok, kinonok), amelyek a baktériumok membránjában, illetve a cianobaktériumok kromatofóráiban is megtalálhatók.
Az utóbbi években azonban vita bontakozott ki a folyamat részleteit és a résztvevő partnereket illetőleg, miután kiderült, hogy az eredeti Margulis-féle elképzelés sarkkövei támadhatóvá váltak. Ezen sarkkövek közül az egyik legjelentősebb, hogy a szimbiózis egy prokarióta és egy „primitív eukarióta” sejt között ment végbe, melyben a sejtfal elvesztése és a fagocitózis voltak a kulcselemek, tehát létezett egy normális sejtmaggal és maghártyával rendelkező, de mitokondrium-mentes ún, “primitív eukarióta” sejt.

1. ábra
Az ábrán jól látszik, hogy ez a feltételezett primitív sejt teljes maghártyával és részleges endoplazmatikus retikulummal is rendelkezett még a mitokondrium ősének érkezése előtt. Ebből alakult ki az élővilág származtatásának korábbi modellje.

2. ábra
Az ábra szerint tehát volt egy jelentős időintervallum az evolúcióban, amely során léteztek “ősi anaerob eukarióták”, azaz olyan sejtmaggal, maghártyával és belső membránrendszerrel rendelkező sejtek, amelyekben nem voltak mitokondriumok. Mindez egybecseng a Föld ősi légkörére vonatkozó korábbi elképzeléssel, az “őslégkör teóriával”, mely szerint a korai időszakban a Föld ősi atmoszférája redukáló volt, amelyben dominált az ammónia (NH3), metán (CH4) és a szén-dioxid (CO2), hiányzott viszont belőle az oxigén (O2). Ezt az elképzelést az 1910-es években alkotta meg egymástól függetlenül egy orosz (Oparin) és egy brit (Haldane) tudós. Mindketten a Naprendszer legnagyobb, és egyik légkörrel rendelkező bolygója, a Jupiter légkörében levő gázok összetételét vizsgálták olyan távcsövekkel, amelyekkel színképelemzési módszerekkel a gázok elemzését és azonosítását el lehetett végezni. Mivel ott a légkörben főleg az említett gázokat találták, azt posztulálták, hogy az ősi Föld légköre is hasonló összetételű volt. Értelemszerű, hogy az első elméletek a korai sejtek energianyerésével kapcsolatban abból indultak ki, hogy mivel az őslégkör redukáló volt, nem volt oxigén a sejtek közelében, ezért azok kizárólag anaerob módon, fermentációval nyerték az energiát, azaz termelték az ATP-t, és ez a típusú energianyerés volt az ATP-gyártás domináns módja egészen addig, amíg az első cianobaktériumok meg nem jelentek és az oxigént termelő fotoszintetikus ATP-szintézist meg nem valósították. Tehát az endoaszimbionta hipotézis szerint a 2.ábrán látható „ősi anaerob eukarióták”-ból alakultak ki a késöbbi eukarióta sejtek, először a mitokondriummal rendelkező állati és gombasejtek, majd a másodlagos endoszimbiózissal az eukarióta sejt által bekebelezett cianobaktériumok rérvén a kloroplasztisszal rendelkező növénysejtek. Ez az elképzelés igen komoly viták forrásává vált az utóbbi időben.
A másik igen sokat hangoztatott premissza (a szimbiózis hajtóereje az oxidatív foszforiláció révén keletkező energiatöbblet volt) is az ellenvélemények kereszttüzébe került. Az újabb molekuláris kutatások ugyanis nem adtak alapot a mitokondrium-nélküli „primitív eukariótsa” sejt létezésének, mivelhogy minden jelenleg vizsgát eukarióta sejtek nukleáris genomjában (azokban is, amelyekben másodlagosan elvesztették a mitokondriumaikat) ki lehet mutatni mitokondriális géneket.. Kiderült továbbá, hogy a fermentatív energianyerés egyáltalán nem ősibb és primitívebb, mint a kemiozmotikus ATP-szintézis.
Az eredeti Margulis-féle elképzelés értelmében a kiindulási sejt valamilyen okból elvesztette sejtfalát. Ennek a fontos eseménynek azonban sem a miértjét, sem a hogyanját nem tudjuk meg az elképzelésből. A sejtfal igen fontos celluláris struktúra, amely többek között ozmotikusan is stabilizálja a sejtet és megvédi az esetleges ozmotikus stresszhatások következményeitől. A sejtfal elvesztése tehát igen komoly válaszút elé kellett, hogy állítsa a sejtet, pl. alternatív ozmoregulációs mechanizmusok (lüktető űröcske) kialakítása felé, amelyről azonban a teória nem tesz említést. Kövessük tovább az endoszimbionta hipotézis logikai menetét. A sejtfal nélküli „primitív eukarióta” sejtben a plazmamembrán betüremkedése következett be és a betüremkedett membránszakaszok belül lefűződve intracelluláris kompartmenteket hoztak létre. (3. ábra).

3. ábra
Előfordulhatott, hogy egy ilyen kompartment körülzárta a cirkuláris DNS-molekula egyik kópiáját, és így alakult ki a sejtmag. Hasonló kompartmentáció eredményezhette a lizoszómák kialakulását. Később a kompartmentek membránjaiból fokozatosan létrejött a belső membránrendszer és a primitív citoszkeletális apparátus fokozatos átalakulásával létrejöttek az eukarióta sejtekre jellemző citoszkeletális struktúrák (4. ábra).
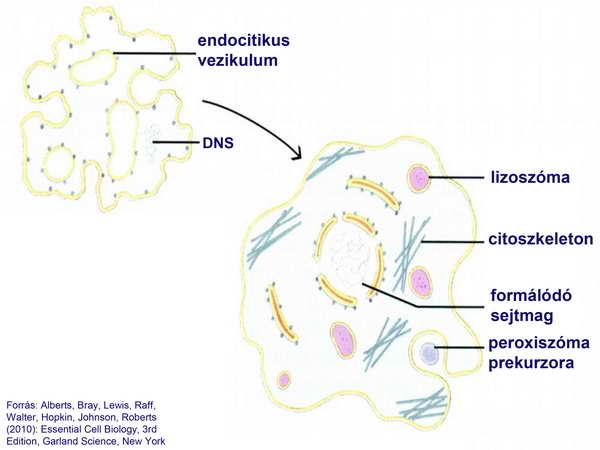
4. ábra
Figyeljük meg, hogy a fokozatosan bonyolódó sejtben már megjelennek a citoszkeletális elemek, sőt megjelenik a peroxiszóma őse is, tehát a sejttérfogat folyamatosan nő, anélkül., hogy annak energetikai hátteréről valamilyen mechanizmussal tudna a sejt gondoskodni. A sejt alapvetően fagocitózissal táplálkozott és a környezetében élő baktériumokat fogyasztotta.. Ezt követően egy kedvező pillanatban ez a primitív eukarióta sejt bekebelezett egy szabadon élő baktériumot, amely szimbiontként együtt maradt a gazdasejttel, sőt abban tovább osztódott és így jöttek a mitokondriumok (5-6. ábrák).
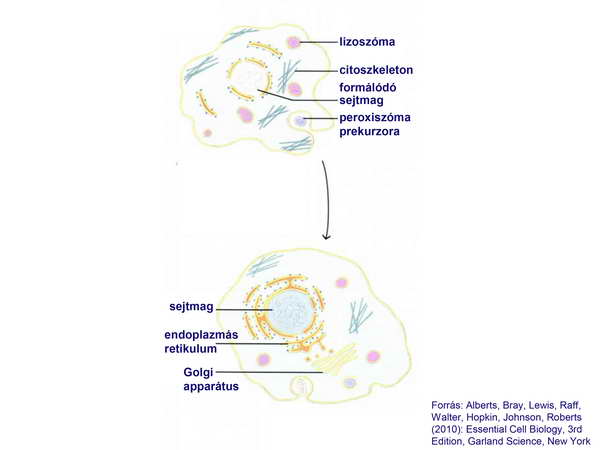
5. ábra

6. ábra
Ezzel új típusú (eukarióta) sejt-organizáció alakult ki, amely sikeresebbnek bizonyult, mint a korábbi prokarióta sejt, ebből az új sejt-típusból alakult ki a fosszilis és ma élő valamennyi állat-, növény- és gombasejt, sőt az egysejtű eukarióták is. A mitokondrium érkezésével a folyamat nem zárult le, cianobaktériumok bekebelezésével (szekunder endoszimbiózis) jöttek létre a növénysejtek (7. ábra).

7. ábra
Az eredeti hipotézissel kapcsolatban azonban problémák is vannak:
A jelenleg ismert összes eukarióta sejt (még azok is, amelyek másodlagosan elvesztették a mitokondriumokat) nukleáris genomjában megtalálhatók a mitokondriális fehérjéket kódoló gének, tehát valamikor minden eukarióta sejt kellett, hogy tartalmazzon mitokondriumokat, azaz nem létezett mitokondrium-nélküli eukarióta sejt. Ez ellentmond az endoszimbionta hipotézis legfontosabb premisszájának, azaz annak, hogy a szimbiózis egy prokarióta és egy „primitív eukarióta” sejt között ment végbe.
NNem teszi világossá, hogy a „primitív eukarióta” sejt kialakulása és belső állománya komplexitásának fokozatos növekedése energetikailag hogyan volt lehetséges.
Azt tételezi fel, hogy az első sejt energianyerése fermentatív alapú volt, és ehhez a mitokondrium által szolgáltatott oxidatív folyamatok adták a szimbiózis hajtóerejét. Mint azt az “Az első közös sejt (LUCA) jellemzése” című fejezetben taglaljuk, ez a feltételezés nem állta ki az idők próbáját.
E problémák megválaszolása és valószínűbb forgatókönyvek felállítása céljából több elméletet is kialakítottak a kutatók, közülük az egyik legnagyobb horderejű a William Martin és a magyar származású Müller Miklós által kifejlesztett hidrogén-hipotézis, amelyet 1998-ban publikáltak a „Nature”-ben. A hipotézis neve onnan származik, hogy szerintük az eukarióta sejt kialakulásának hajtóereje a metabolikus szintrófia (metabolikus egymásra utaltság, a partnerek felhasználják egymás anyagcsere-termékeit), melyben nem az oxigén, hanem a hidrogén játszotta a vezető szerepet. Az elmélet abból indul ki, hogy egy térfogatában jelentősen megnőtt eukarióta sejtnek igen magas az energia-igénye, hiszen kompenzálnia kell a csökkenő felszín/térfogat arányt, tehát az eukarióta sejt kialakulása és az energiát szolgáltató mitokondrium ősének bekebelezése szimultán kellett, hogy végbemenjen. Más szóval minden érv ellentmond annak, hogy mitokondrium-nélküli „primitív” eukarióták léteztek volna. A hipotézis kifejlesztéséhez az első lökést az archaebaktériumok felfedezése volt 1977-ben. Ezek a prokarióták extrém körülmények között élnek (hőforrások, extrém sós környezetek), amelyekről korábban az volt az álláspont, hogy az élettel összeegyeztethetetlenek. Felfedezésük után kiderült, hogy az Archaeák sejtfelépítésüket, metabolizmusukat és enzimatikus apparátusukat tekintve igen kevés rokonságot mutatnak az addig „Baktériumok”-nak nevezett csoporttal, amelyeket azóta Eubaktériumoknak neveznek. Néhány lényeges különbséget az alábbiakban foglalunk össze:
az Archaeák membránjában speciális lipidek találhatók, amelyek három vonatkozásban is különböznek az Eubaktériumok membránjaiban található lipidektől:
a/ Az Archaeákban éter-lipidek találhatók, azaz olyan lipidek, amelyekben a glicerin két szénatomja éterkötéssel (R-O-R) kapcsolódik egy-egy alkillánchoz, szemben az Eubaktériumokkal, ahol észter-kötéssel kapcsolódó lipidek vannak jelen (tehát amelyekben a glicerin két OH-csoportja és egy-egy zsírsav COOH-csoportja között vízkilépéssel észterkötés jön létre),
b/ Az Archaeákban a két alkillánc a glicerin sn-2 és sn-3 pozíciójához kapcsolódik, szemben az Eubaktériumokkal, ahol a két zsírsavlánc az sn-1 és sn-2 pozíciókat észteresíti. Emiatt a glicerin-foszfát csoport sztereokémiája pontosan fordítottja egymásnak a két baktérium-csoportban (enantiomerek),
c/ az Archaeákban nem zsírsavak, hanem izoprén-egységekből álló szénhidrogén-láncok kapcsolódnak a glicerinhez (8. ábra).

8. ábra
A legtöbb Archaea sejtfalából hiányzik a peptidoglukán, bennük a sejtfalat felépítő fehérjék egy rigid réteget hoznak létre a sejt körül (pszeudomurein). Mindezek a különbségek hozzájárulhatnak az Archaeák szélsőséges tűrőképességéhez. A legmegdöbbentőbb felfedezés azonban az volt, amikor összehasonlító 16S rRNS-szekvencia vizsgálatokból kiderült, hogy az Archaeák közelebbi rokonságban állnak az Eukariótákkal, mint az Eubaktériumokkal. Az utolsó közös őssejtből (Last Universal Common Ancestor, LUCA), amely minden fosszilis és jelenkori élő szervezet őse, kiinduló fejlődési vonal egy ponton elágazott és egyik irányban az Eubaktériumok haladtak tovább, míg a másik irányban létrejött az Archaeák és az Eukarióták közös őse. Ez feltehetőleg egy metanogén (metánt termelő) Archaebaktérium volt.
A hipotézis további kimunkálásához a szerzők számba vették az eukarióta sejtekben fellehető energianyerő folyamatokat, amelyek a következők:
Aerob körülmények között a sejtekben a mitokondriumok termelik az ATP-t kemiozmotikus úton oxidativ foszforiláció révén.
Azokban a sejtekben, amelyek másodlagosan alkalmazkodtak az anaerob viszonyokhoz, sokszor elvesztek a mitokondriumok. Ezekben a sejtekben nincsenek speciális energia-termelő sejtorganellumok, hanem a citoplazmában fermentáció (szubsztrát-szintű foszforiláció) megy végbe.
Vannak olyan anaerob körülmények között élő sejtek, amelyek rendelkeznek hidrogenoszómával, bennűk hidrogén-termeléshez kapcsolt ATP-termelés megy végbe. A hidrogenoszómát New Yorkban élő honfitársunk, Müller Miklós fedezte fel és publikálta 1978-ban. Egyes parazitikus eukarióta egysejtűek (Trichomonas, Guardia, Entamoeba) kettős membránnal határolt, ATP-termelő organelluma, amely piruvátból H2-t és CO2-t termel anaerob körülmények között. A piruvát oxidatív dekarboxilezését a piruvát:ferredoxin oxidoreduktáz (PFO) végzi egy ferredoxin-igényes folyamatban. A redukált ferredoxin oxidálását a hidrogenáz végzi, amely két protonból H2-t állít elő. A PFO működéseként termelődő acetil-CoA-t az acetát:szukcinát CoA szintetáz alakítja tovább acetáttá és szukcinil CoA-vá, melyből a szukcinil CoA szintetáz működésének eredményeként keletkezik az ATP.

9. ábra
A hidrogenoszómák felfedezését követően Müller Miklós azt a lényeges észrevételt tette, hogy azok közvetlen közelében baktériumok találhatók az anaerob protozoa sejt belsejében. Ezekről később kiderült, hogy metanogén Archaeák.
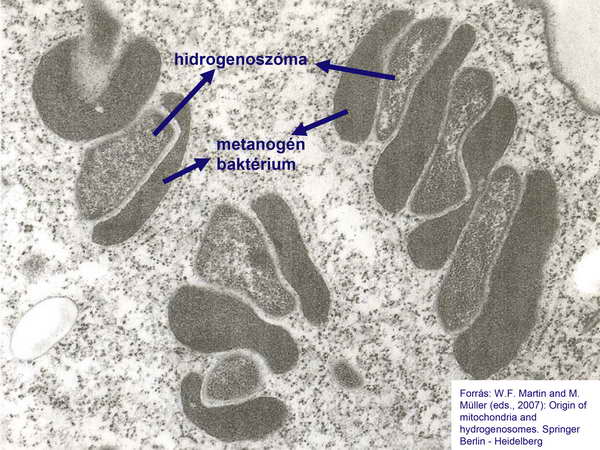
10. ábra
Ez a közös előfordulás nem lehet véletlen, és a közöttük feltételezett anyagcsere-termékek közvetítette kapcsolat adta az első lökést a hidrogén-hipotézis kifejlesztéséhez. A hidrogenoszóma ugyanis pontosan azokat az anyagokat termeli, amelyeket a metanogén Archaebaktérium fogyaszt (H2 és CO2).
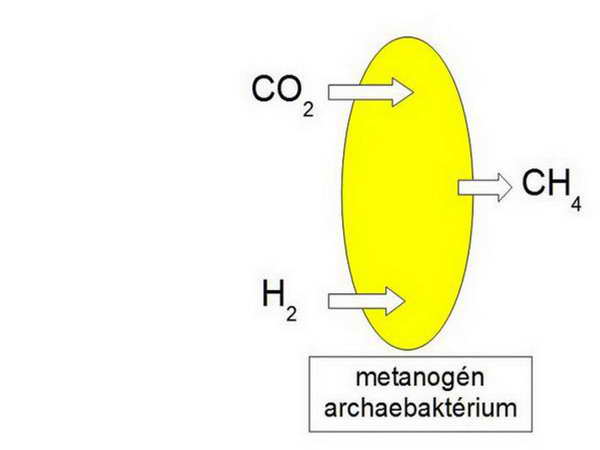
11. ábra
Tehát, ha a szimbiózis él és működik egy anaerob protozoa belsejében ma, miért ne működhetett volna 2 milliárd évvel korábban is? Hiszen ez megmagyarázná, hogy miért volt a két sejt egymásra utalva és egy teljesen új értelmezést adna az eukarióta sejt kialakulásához szükséges szimbiózis mikéntjének.
A hidrogén-hipotézis szerint az eukarióta sejtek kialakulásának kulcsmomentuma két prokarióta sejt között vegbemenő szimbiózis volt. A „befogadó” partner (gazdasejt) egy metanogén archaebaktérium volt, amely anyagcseréjét tekintve autotróf volt, a „jövevény” (szimbiont) pedig egy fakultatív anaerob (az angol eredetiben: versatile) eubaktérium volt, anyagcseréjét tekintve heterotróf. A metanogén Archaebaktérium egyrészt 16S rRNS-beli közelsége, másrészt speciális energianyerési mechanizmusa miatt került a képbe. A metanogén baktériumok szén-dioxidot használnak elektron-forrásként, és hidrogént használnak elektron-donorként, azaz tulajdonképpen a szén-dioxidot redukálják metánná. E többlépéses folyamat kezdeti lépései a citoplazmában, utolsó lépése viszont a membránban megy végbe, ahol a H atomokból származó elektronok kerülnek rá a végső elektron-akceptorra, a felszabaduló protonok pedig átpumpálódnak a membránon, létrehozva az ATP-szintézishez szükséges kemiozmotikus proton elektrokémiai potenciál-különbséget. Az ATP birtokában felépítik saját szén-vegyületeiket (autotrófok), közöttük szénhidrátokat is.
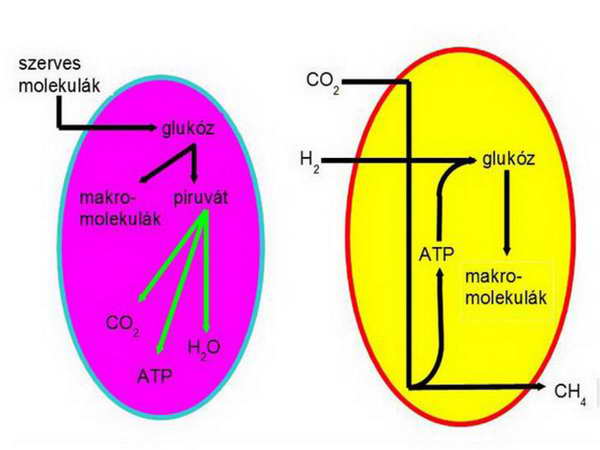
12. ábra
A hidrogén-hipotézis alapján két olyan baktérium találkozott, amelyek között metabolikus függőség bontakozott ki. Az elképzelés szerint a két baktérium anaerob környezetben találkozott egymással, és egy darabig egymás mellett, de egymást nem zavarva éltek. A metanogén egy darabig a geológiai forrásból származó H2-t és CO2-t „fogyasztotta”, és metánt termelt a környezetének, az eubaktérium pedig (amely legnagyobb valószínúséggel egy α-proteobaktérium lehetett) a környezetéből felvett szerves anyagokat vagy glukózt lebontva egyrészt szubsztrát szintű ATP-szintézist végzett, másrészt a piruvátot hidrogenoszómaként bontotta le (azaz H2–t és CO2-t termelt és azokat kibocsátotta a környezetébe), és így további ATP-hez jutott (12. ábra). A geológiai források csökkenésével a metanogén egyre inkább „rászorult” egy, a közelébe került eubaktérium termékeire, és így lassanként egyre szorosabb lett a metabolikus függőség, anaerob szintrófia jött létre.
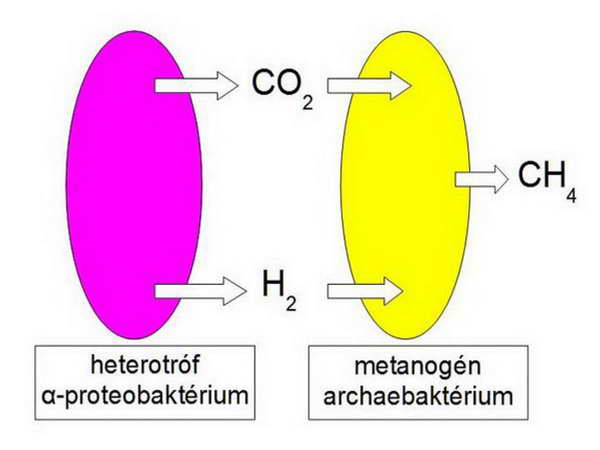
13. ábra
Idővel a metanogén ráutaltsága a baktérium által termelt termékekre egyre erősebb lett, és az archaebaktérium egyre közelebb került az eubaktériumhoz, és egy adott pillanatban bekerült annak belsejébe. Az internalizációs lépés a további jelentős eltérés az endoszimbionta hipotézissel szemben. A hidrogén-hipotézis szerint ugyanis nem volt fagocitózis, ugyanis mindkét baktérium partnernek megvolt a saját sejtfala. Szerintük feltehetőleg a szimbiont termelhetett sejtfallazító, vagy -bontó enzimeket, amivel könnyen elérhette, hogy a metanogén baktérium sejtfala lokálisan annyira fellazult, hogy a szimbiont át tudott jutni a sejtfalon. A gazdasejt sejtmembránja azonban teljes egészében körülvette a bekerült szimbiontot, amely tehát egy kettős membránnal körülvett sejtrészecskévé vált.
Ezután a viszonyok fokozatosan stabilizálódtak és a következő, mindent eldöntő lépés a horizontális gén-transzfer volt, amelynek során a szimbiontból gének kerültek át a gazdasejtbe. A teória szerint először a glukóz felvételében és lebontásában (glikolízis) szerepet játszó enzimek génjei kerülhettek át, amelyek teljesen átalakították a gazdasejt metabolizmusát (14. ábra).

14. ábra
A gazdasejt fokozatosan megszűnt metánt termelni, mert anyagcseréje átállt a glukóz fermentatív lebontásából történő energia-nyerésre, és a képződő piruvátot visszajuttatta a szimbiontnak.
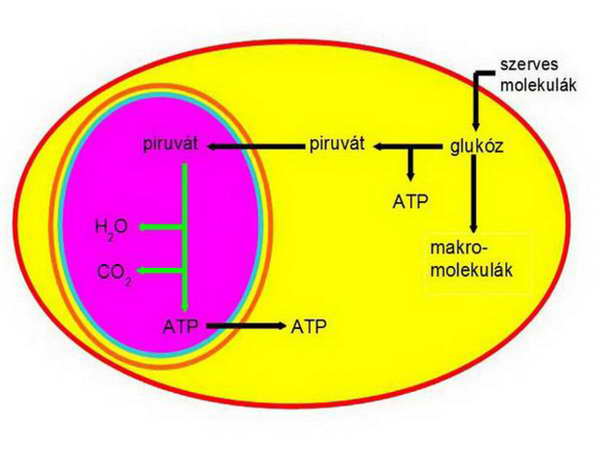
15. ábra
Ezt a szimbiont kezdetben fermentatív úton bontott le, de a termelődött ATP-t már a gazdasejt rendelkezésére bocsátotta. Ezt követően azonban a szimbiont fokozatosan átállt oxidatív anyagcserére, és anyagcseréje a kezdeti, hidrogenoszómára jellemző ATP-termelésből fokozatosan átállt mitokondrium-szerű ATP-termelésre.

16. ábra
Ezzel kialakult a gazdasejt és a szimbiont bensőséges kapcsolata és egy minőségileg új rendszer jött létre. Később a szimbiont osztódásával az új sejt egyre több energiát állított elő, ezzel megvalósulhatott a sejtméret növekedése, ami az Eukarióta sejtek kialakulásának következő fontos lépése volt. Ez az új sejttípus most már elmozdulhatott a szigorúan anaerob környezetből, és olyan területeket is meglátogathatott, ahol jelen volt az oxigén. Az oxidatív energianyerés révén több ATP-t tudott gyártani, ami jelentős szelekciós előnyhöz juttatta, azaz az új sejttípus egyre versenyképesebb lett.
A sejtmag kialakulása. A glikolízis enzimeket kódoló gének átkerülése után idővel további gének is átkerültek a korábbi metanogén (most már gazdasejt) genomjába. Ilyenek voltak, pl. a lipid-bioszintézis enzimeit kódoló gének. A gazdasejtben szintetizálódó eubakteriális (észter-típusú) lipidek elkészülte után azonban feltehetőleg elveszett a „rendeltetési hely”-re vonatkozó utasítás, így membránalkotó eubakteriális lipidek szintetizálódtak anélkül, hogy rendeltetési helyük a sejt számára pontosítva lett volna. A citoplazmában fokozatosan felhalmozódó lipidek tehát spontán asszociálódtak egymással, először micellákat, majd liposzómákat, vezikulumokat és végül kettős lipidrétegekből szerveződő membránokat hoztak létre a sejten belül. Ezek a membránok körülvehették a cirkuláris DNS egyik kópiáját, és így keletkezhetett a membránnal körülhatárolt sejtmag, és így jöhetett létre a a maghártyával összefüggő belső membránrendszer is.
Az eukarióta plazmamembrán kialakulása. A gazdasejt (korábbi metanogén) membránjában éter-lipidek domináltak, amelyek lehetővé teszik, hogy az Archaebaktériumok meg tudjanak telepedni extrém szélsőséges körülmények között is. Ezekben a lipidekben azonban nem zsírsavak, hanem poli-izoprén szénhidrogén-láncok fordulnak elő, így a belőlük felépülő membránokban nincs lehetőség a membránfluiditás szabályozására. Az új (immár eukarióta) sejt viszont könnyen kerülhetett olyan körülmények közé, ahol az erre való képesség szükséges lehetett (pl. hidegebb körülmények közé). Ekkor következhetett be az a fordulat, amelynek során eubakteriális (észter-típusú) lipidek váltották fel az éter-típusú lipideket és létrejött az eukarióta sejt plazmamembránja. Azt, hogy az ősi izoprén-tartalmú lipidek már az első eukariótákban jelen voltak, az bizonyítja, hogy minden jelenkori eukarióta sejtben és egyedben megvannak az izoprén-bioszintézis génjei.
Összefoglalva, a hidrogén-hipotézis abból a megfigyelésből indul ki, hogy mitokondrium nélküli eukarióta sejt nem létezett, az eukarióta sejt kialakulásának kulcslépése tehát a mitokondrium létrejötte és nem a maghártyával körülhatárolt sejtmag kialakulása volt. A szimbiózis két prokarióta sejt között ment végbe, annak hajtóereje, pedig metabolikus szintrófia volt, melyben a hidrogén játszotta a kulcsszerepet. A szimbiózis eredményeként létrejött új szerveződési egység (eukarióta sejt) metabolikus viszonyait, valamint a belső kompartmentek kialakulását a szimbiontból a gazdasejt felé irányuló horizontális gén-transzfer tette lehetővé.
Mióta vannak eukarióta sejtek a Földön?
Mik az endoszimbionta hipotézis pozitív vonatkozásai?
Mi az endoszimbionta hipotézis legnagyobb hibája?
Miért nem mitózis a baktériumok osztódása?
Ki és milyen sejtekben fedezte fel a hidrogenoszómákat?
Milyen anyagcserét folytat és hogyan termel ATP-t a hidrogenoszóma?
Milyen reakciót katalizál a PFO?
Milyen megfigyelés szolgáltatta az alapot a hidrogén-hipotézis kifejlesztéséhez?
Milyen anyagcserét folytat és hogyan termel ATP-t egy metanogén Archaebaktérium?
Mit jelent a „metabolikus szintrófia”?
Mi a horizontális és mi a vertikális gén-transzfer?
Mely gének átkerülése állította át a metanogén gazdasejt anyagcseréjét a hidrogén-hipotézis szerint?
Mely gének átkerülése járult hozzá a maghártya kialakulásához a hidrogén-hipotézis szerint?
Hogyan alakult ki az eukarióta sejtmembrán a hidrogén-hipotézis szerint?
Mi a hidrogén-hipotézis jelentősége?