Tartalom
Az eukarióta sejt kialakulásáról szóló fejezetben láttuk, hogy az eukarióta sejt újszerűsége nem annyira a kromatinállomány és a sejtmaghártya létrejöttében rejlik (mint azt a Margulis-féle klasszikus endoszimbionta hipotézis hirdeti), hanem sokkal inkább abban a tényben, hogy az eukarióta sejt egy olyan minőségileg új formáció, amelyben több különböző genommal és fehérjeszintetizáló apparátussal rendelkező (tehát elvileg autonóm) rendszer alkot egy közös, tehát összehangoltan működő egységet. Ez azért egy roppant izgalmas kérdés, mert az autonómia és a kooperáció elvileg egymást kölcsönösen kizáró kategóriák. Az eukarióta sejt mégis megvalósitja a látszólag lehetetlent, mert hierarchikus rendet alakít ki a sejten belüli genetikai állományok között. Fokozatosan növekvő fontosságra tesz szert a gazdasejt genomja (amit tehát mostantól nukleáris genomnak hívunk), az organellumok pedig fokozatosan veszítenek önállóságukból. Mindennek a hátterében egy meghatározó és mindent eldöntő mozzanat áll: a horizontális gén-transzfer.
A horizontális gén-transzfer fogalma szélesebb léptékben is értelmezhető, mint sejten belüli viszonyok között, és általánosságban azt a jelenséget értjük alatta, amikor egy időben egymás szomszédságában létező objektumok között valósul meg a genetikai anyag átadása. Összehasonlításként: vertikális gén-transzfernek a generációk közötti (szülőtől az utód felé történő) genetikai állomány átadást értjük.
Az evolúció korai szakaszában a horizontális gén-transzfer mind sejtek, mind a sejten belüli genommal rendelkező organellumok között egyaránt lényeges, gyakori és uralkodó mozzanat volt. Sőt, olyan elképesztő mértéket ért el, hogy pl. emberben a mitokondriális DNS mindössze 13 fehérjét kódoló gént tartalmaz, a több ezer további mitokondriális fehérjét kódoló gén viszont átkerült a sejtmagba (azaz a szimbiózis korábbi gazdasejtjének genomjába). Hasonló módon, a cianobaktériumokban található mintegy 3000 fehérjéből mindössze 100-150 fehérjét kódoló gén maradt meg a jelenlegi kloroplasztiszokban, a többi átkerült a növénysejtek nukleáris genomjába. Mindennek egyenes következménye, hogy az organellumok gyakorlatilag feladták az önálló létezés lehetőségét, így sem a mitokondriumok, sem a kloroplasztiszok nem képesek önállóan (a sejten kívül) életben maradni, hiszen a bennük található fehérjék többsége nukleáris kódolású és citoplazmikus transzlálódású lett. Ha ez a gén-áramlás ilyen komoly méreteket öltött, felmerül a kérdés, hogy vajon miért maradtak meg egyáltalán gének az organelláris genomokban és miért nem került át az összes gén, hiszen ezért a maroknyi gén által kódolt üzenetért önálló transzkripciós és transzlációs apparátust működtetni nem tűnik evolúciós szempontból ésszerűnek, tehát valami fontos oka kell, hogy legyen annak, hogy ez a néhány gén megmaradt eredeti helyén. Erre a fontos kérdésre többen is megkíséreltek választ adni, közülük a legelfogadottabb hipotézist John F. Allen publikálta 1993-ban a Journal of Theoretical Biology című folyóiratban „CO-location of Redox Regulation” címmel, amit azóta CORR hipotézisnek neveznek és magyarul kb. „Helyszínen végzett redox szabályozást” jelent.
Abból a korábbi megfigyelésből indult ki, hogy a redox-potenciál képes a génexpresszió szabályozására. A szabályozást a bakteriális rendszerekből jól ismert két-komponensű rendszer végzi, amelynek egyik komponense (szignál szenzor kináz) az elektrontranszport-lánc közvetlen közelében, míg a másik komponens (válasz regulátor) a mitokondrium, vagy kloroplasztisz genomban az illető fehérjét kódoló gén promótere előtt helyezkedik el (1. ábra).
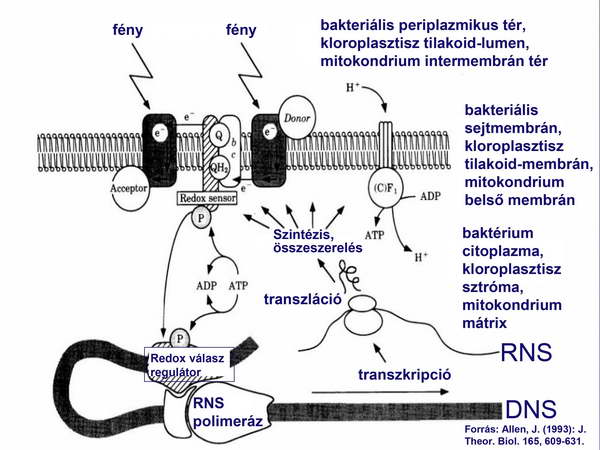
1.ábra
A szenzor kináz autofoszforlálja saját magát egy hisztidin aminosavon (2. ábra),
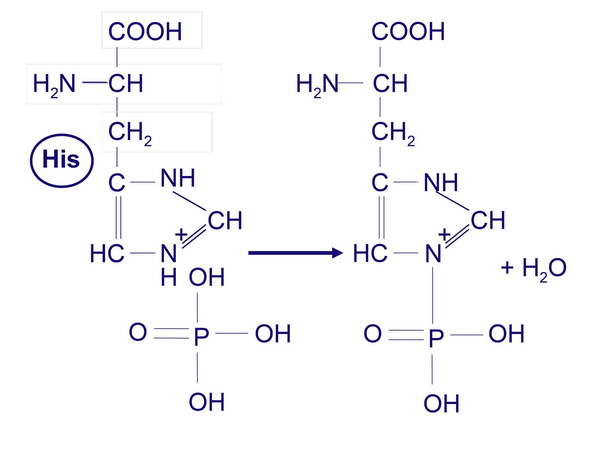
2. ábra
majd a foszfát átkerül a válasz-regulátor aszparaginsav-csoportjára (3. ábra).
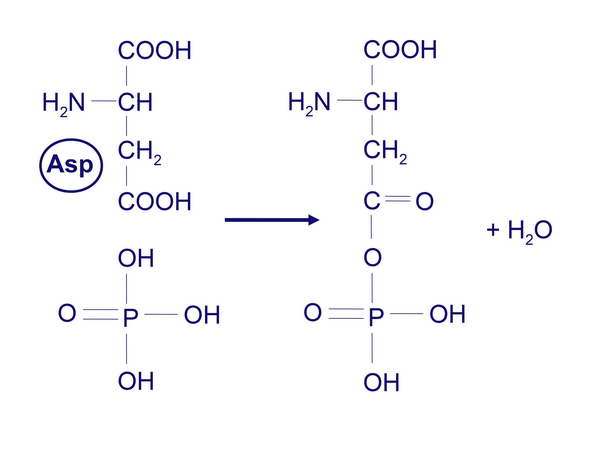
3. ábra
Mind a mitokondrium, mind a kloroplasztisz energiatermelő organellumok, amelyek elsőrendű feladata az ATP előállítása. Az ATP szintézis leghatékonyabban kemiozmotikus úton valósul meg, ehhez viszont elektronátmeneti energiára van szükség. A fejlettebb rendszerekben az elektronátmeneti energia olyan tetemes, hogy az elektron-donor és –akceptor nem közvetlenül reagál egymással, hanem köztük további komponensek találhatók, amelyeket elektrontranszport-lánc tagoknak is nevezünk. Mindkét organellumban az elektrontranszport sebessége a legfontosabb a légzés, illetve a fotoszintézis hatékony lebonyolítása, optimalizálása és minél több ATP termelése szempontjából. Előfordulhat, hogy valamelyik elektrontranszport-lánc komponens a működés közben meghibásodik, így miatta az egész elektrontranszport lelassul és az ATP-szintézis hatékonysága romlik. Ilyenkor gyors beavatkozásra van szükség, amihez a leghatékonyabb, ha helyi (lokális) döntések meghozatalára és megvalósítására kerül sor. Ezeket a döntéseket és helyreigazításokat végző struktúráknak tehát ott kell lenniük a hiba közvetlen közelében, hogy gyorsan és hatékonyan ki tudják azt javítani. Az ilyen komponenseket kódoló gének tehát nem kerültek át a sejtmagba, hanem megmaradtak az organelláris genomokban.
Allen professzor felvázolja a horizontális gén-transzfer feltételezett folyamatát is:
Kezdetben valamennyi organelláris (mitokondrium, illetve kloroplasztisz) fehérjét az organelláris genom kódolt, és azok organelláris riboszómákon szintetizálódtak. Közülük bizonyos kitüntetett gének transzkripciója redox szabályozás alatt állt, mert az általuk kódolt fehérje az elektrontranszport-lánc egyik fontos komponense volt (4. ábra).
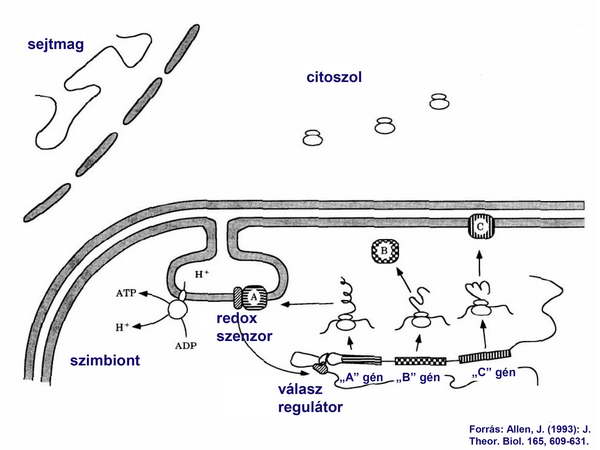
4. ábra
Később valamennyi organelláris fehérjét kódoló gén átkerült a nukleáris genomba is, de megmaradt az organelláris genomokban is. Így minden organelláris fehérje elkészült citoplazmikus, illetve organelláris riboszómákon is. Ezen fehérjék közül a nem redox-regulált fehérjék nukleáris kódolása és citoplazmikus transzlációja volt az előnyösebb alternatíva, főleg a nukleáris genom nagyobb védettsége (hisztonok, repair rendszerek) miatt, így ezek a gének fokozatosan elvesztek az organelláris genomokból (5. ábra).
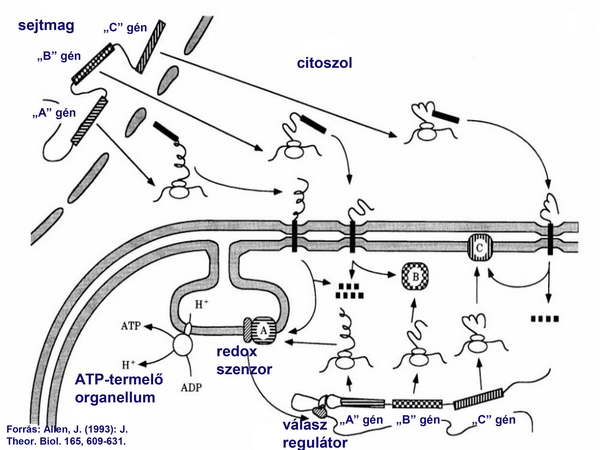
5. ábra
3/ A redox-regulált gének esetén viszont a helyi organelláris kódolás bizonyult az előnyösebb alternatívának, mert így az elektrontranszport sebességét közvetlenül lehet szabályozni. Emiatt ezen gének nukleáris változataira már nem volt szükség, tehát azok fokozatosan elvesztek a nukleáris genomból (6. ábra).
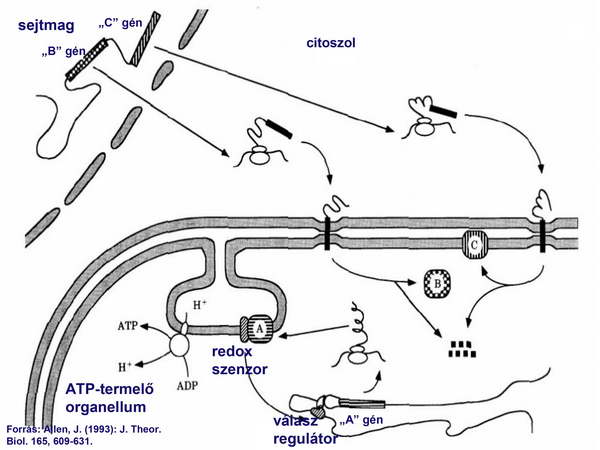
6. ábra
A hisztidin foszforilációja
Az aszparaginsav foszforilációja