Tartalom
Az eukarióta sejt belsejében, a prokariótákkal szemben, sokféle, különböző funkciójú membrán-határolta teret –kompartimentumot – találunk. Kompartimentumokként jelenik meg a sejtszervecskék többsége (sejtmag, mitokondrium, stb.) Ezeknek a belső tereknek a kialakulása tette lehetővé, hogy az eukarióta sejtekben többféle, eltérő környezetet igénylő, sokszor egymást kizáró metabolikus folyamat tud egyidőben működni. Mindezek mellett az organellumok megjelenése a sejt belső felszínének radikális növekedésével hozzájárult az alapvető membránhoz kötött biokémiai folyamatok (például az oxidatív foszforiláció) hatékonyságának emelkedéséhez.
A kompartimentumok megjelenése a vitathatatlan előnyök mellett azonban nagy kihívást is jelent a sejt számára. Ismert, hogy valamennyi fehérje a citoplazmában található riboszómákon szintetizálódik. A sejtnek tehát gondoskodnia kell valahogyan arról, hogy azok, de csakis azok a fehérjék, amelyek egy adott térrészben működnek, el is jussanak rendeltetési helyükre. Emellett minden sejt folyamatosan juttat a sejten kívüli extracelluláris térbe (vagy a plazmamembránba) olyan fehérjéket, amelyek például a szomszédos sejtekkel vagy a sejten kívüli állománnyal való összeköttetést és a kommunikációt biztosítják. Ez különösen igaz az úgynevezett szekréciós sejtekre, amelyek a soksejtű szervezet váladéktermelésre szakosodott sejtjei.
Hogyan oldható meg ez a bonyolult feladat? Milyen természetű az a jel, amely „megmondja”, hogy melyik fehérje hová kerüljön a sejtben? Milyen mechanizmus valósítja meg az „utazást” a célállomásra? A kérdéseket megválaszoló munkáért Günter Blöbel német származású amerikai kutató 1999-ben Nobel-díjat kapott. Szignál-elmélete szerint: 1) a fehérje integráns részét képező szignálpeptidek biztosítják a megfelelő sejtszervecskébe jutást; 2) létezik olyan szállítóapparátus, amely felismeri ezt a jelet, és biztosítja a célbaérést.
Azt, hogy egy fehérje milyen módon kerülhet rendeltetési helyére, meghatározza, hogy a szintézis helyének tere (a citoszol) egyenértékű-e, ekvivalens-e a célszervecske terével. Ebből a szempontból meg lehet különböztetni topológiailag ekvivalens tereket (például citoszol – sejtmag, endoplazmás retikulum belső tere – Golgi-készülék belső tere), illetve nem-ekvivalens tereket (citoszol – endoplazmás retikulum tere, citoszol – mitokondrium) (1. ábra). Az egymással egyenértékű közegek közös evolúciós eredetűek (2. ábra), közöttük szabadabb az átjárás. Ezt jelzi a nagyméretű vizes pórus a sejtmag esetében, illetve a vezikuláris transzport a szekréciós és endocitotikus útvonalon belül. Nem-ekvivalens terek között csak szigorúan szabályozott, zárható csatornákon, transzlokátorokon keresztül történhet a fehérje átjutása, hiszen egyébként sérülne az eltérő terek integritása.
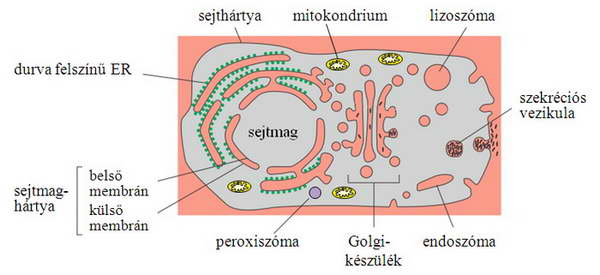
1. ábra. Különböző belső terek az állati sejtben. A topológiailag ekvivalens terek azonos színnel vannak jelölve.
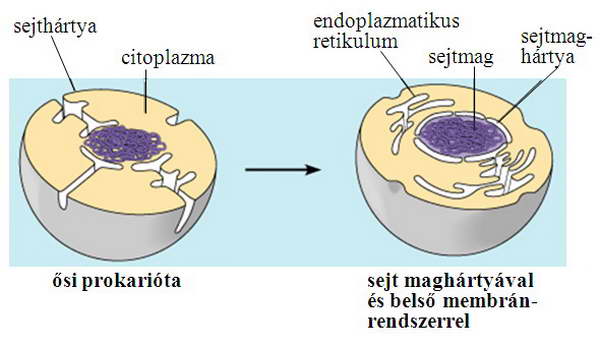
2. ábra. A sejt belső membránrendszerének evolúciós eredete