Az eukariota sejtek képesek alak- és helyváltoztatásra. A mozgásjelenségek hátterében a vázrendszer – elsősorban az aktinváz és a hozzákapcsolt motorfehérjék – aktivitása/átrendeződése áll. A mozgásjelenségeket legegyszerűbb hordozó felületre kitelepített mozgékony sejteken – neutrofil leukocitákon, fibroblasztokon, amöboid sejteken - vizsgálni. Alapállapotban ezeken véletlenszerű irányokban folyamosan képződő kis, rövid élettartamú kitüremkedések jelennek meg. Különböző anyagokkal pozitív kemotaxis váltható ki: a véletlenszerű álláb képződés leáll, sejt irányított mozgásba kezd, megindul az inger forrása felé, abba az irányba, ahol a kemotaktikus anyagok koncentriója a legnagyobb. A folyamat első lépése a mozgás irányának kijelölése. Ez azért lehetséges, mert a kemotaxist kiváltó anyagokat a plazmamembránba ágyazott receptor molekulák érzékelik. A receptorok egyenletesen oszlanak el a sejt felszínén, közülük azok fognak legerősebben aktiválódni, melyek a sejtnek az ingerforrás felé néző oldalán helyezkedenek el. Az aktivált receptorok jelátviteli folymatokat indítanak be, melyek eredményeként ebben a régióban lokálisan megnő egy enzim, a foszfatidil-inozitol-3kináz (PI3K) aktivitása és felhalmozódik többféle kis G-fehérje. Közöttük találjuk a Rho család két tagját, a Rac-ot és a Cdc42-t. A PI3K foszforilálja a plazmamembránban található foszfatidil-inozitol molekulákat az inozitol gyürű 3-as poziciójában. Az így “kijelölt” membránokhoz számos más fehérje kötődik. A PI3K hatását a sejt hátsó (az ingerforrástól távolabbi) oldalán egy foszfatáz, a PTEN semlegesíti . A hátsó régióban megfigyelhető Rho és miozinII felhalmozódás is. E folyamatok végeredményeként kialakul a polarizált sejt, melynek “eleje” és “vége” az említett molekulák egyenlőtlen eloszlásával jellemezhető (34.ábra).
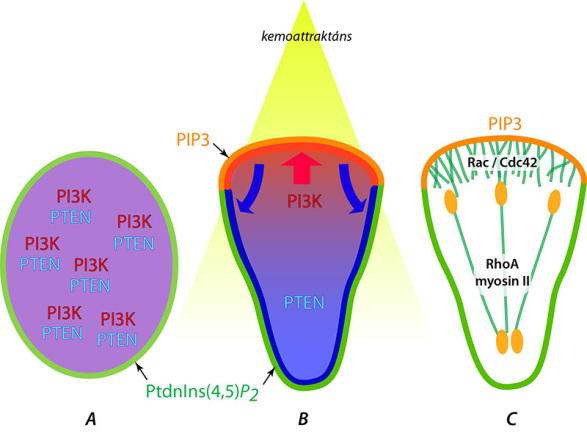
34. ábra Az irányított sejtmozgás lépései és néhány jellemző marker fehérje lokalizációjának változásai. (A) Alapállapotban (ingermentes közegben) a PI3K és a hatását ellensúlyozó foszfatáz, a PTEN, homogén eloszlású, ezért a plazmamembránban nincs PIP3 (PtdIns(3,4,5)P3), a PIP2 (PtdIns(4,5)P2) eloszlása egyenletes. Kemoattraktáns jelenlétben (B) a sejt polarizálódik, amit az eloszlás megváltozása tükröz: a PI3K a sejt ingerfelőli oldalán akkumulálódik és itt PIP3 képződést katalizál, a PTEN a sejt oldalsó és hátsó részére szorul, ahol gátolja a PIP3 megjelenését a plazmamembránban. (C) A polarizált sejt elülső részén aktiválódik két kis G-fehérje, a Rac és a Cdc42, melyek beindítják a WASP/ARP komplex működését, a kialakuló aktinhálózat előrenyomja a plazmamembránt, a sejt lamellipodiumot bocsát az inger irányába. A mozgó sejten letapadási korongok (narancssárga oválisok) jelennek meg, melyek között aktin kábelek húzódnak és miozin II, valamint a RhoA kis G-fehérje jelenléte is kimutatható. Valószínűleg ez a rendszer húzza előre a mozgó sejt hátsó részét.
A következő lépésben megindul az aktinhálózat átszervezése. A Rac és Cdc42 aktiválja a (már korábban ismertetett) WASP-ARP rendszert, melynek hátására sejt elülső régiójában lokálisan dendritikus aktin szövedék alakul ki . A növekvevő aktinszálak kifelé nyomják a plazmamembránt, a sejt elkezd vékony lemezszerű kitüremkedést, lamellipodiumot képezni az ingerforrás irányába (29.ábra). Ezzel egyidejűleg számos letapadási korongok is kialakul (l. “Adheziós molekulák” fejezet), melyek segítségével a sejt rögzül az aljzathoz . A letapadási korongok között aktinszálakból álló kötegek figyelhetők meg, melyekben miozinII is jelen van. Ez a rendszer összehúzódásra képes és felel a sejt hátsó részének előrehúzásáért. A sejtek irányított mozgása tehát összekapcsolt, ciklikusan ismétlődő eseménysor – polarizálódás, kitüremkedés előre, letapadás, a hátsó rész előrehúzása, új letapadási korongok képzése elöl, a hátsó letapadási pontok szétszerelése – eredményeként jön létre. A kitüremkedést a dendritikus aktinhálózat növekvő szálai tolják előre, ez tehát polimerizáció kiváltotta mozgás. Ezzel szemben a sejttest előrehúzását motorfehérje, a miozinII, végzi. Bár a mechanizmus különböző, a munkavégzéshez szükéges energiát mindkét esetben az ATP hidrolízis szolgáltatja.