Az eukarióta sejtek osztódási ciklusát szoros időrendben lejátszódó és szigorúan szabályozott folyamatok sorozata jellemzi. E folyamatok meghatározott sorrendben visszafordíthatatlan változásokat generálnak a ciklusban lévő sejtekben.
A soksejtűek esetében a sejtek ciklusba lépéséhez nélkülözhetetlen a mitogén hatást biztosító növekedési faktorok jelenléte, melyek jelátviteli útvonalak közvetítésével aktiválják a ciklus beindításához szükséges géneket (szérumfüggőség).
A sejttípusok többsége csak akkor képes osztódni, ha a sejthártya integrin molekulái közvetítésével kapcsolatot tudnak kialakítani az extracelluláris mátrix megfelelő komponenseivel (laminin, fibronektin). Az ún. fokális kontaktusok a fokális adhéziós kinázokat aktiválva, jelátviteli folyamatokat elindítva indukálják a sejtek osztódását (letapadás függőség).
A sejt-sejt kapcsolatok kialakulása (szintén szignalizációs útvonalakon keresztül) a normális működésű sejtekben gátolja a ciklust. Ennek eredményeképpen a sejtek a rendelkezésre álló felületet beborítják, kitöltik, majd az osztódás leáll (denzitásfüggő osztódásgátlás). Ebben szerepe lehet annak is, hogy a rendelkezésre álló növekedési faktorok hozzáférhetősége korlátos.
A soksejtű szervezetekben az osztódások/ciklusok száma limitált. Szövettenyészetekben kimutatható, hogy a normál sejtek esetében a ciklusszám fajra jellemző. Például ember esetében embrionális fibroblasztok szövettenyészetében a sejtek kb. 50 sejtcikluson mehetnek át. Ezt követően G0 fázisba lépnek, mitogén hatásokra már nem reagálnak, majd elpusztulnak. E jelenség hátterében a kromoszómák végdarabja, a telomera ciklusról-ciklusra történő rövidülése áll.
A ciklus meghatározott pontjain molekuláris mechanizmusokon alapuló ún. ellenőrző pontok működnek. Szerepük az, hogy a ciklusban való továbblépést csak az adott szakaszra jellemző részfolyamatok teljesítését követően engedélyezik.
Mutációs eredetű hibák a sejtciklus szabályozásának felborulását eredményezhetik. Kialakulhatnak szérumfüggetlen, letapadás független, akár korlátlanul osztódó, ún. immortális sejtek/sejtvonalak. A szabályozás felfüggesztése a sejtek kontrolálatlan osztódásához, sejtburjánzás kialakulásához vezethet.
Az egysejtű és többsejtű eukarióta szervezeteket, illetve a soksejtűek különböző sejtjeit vizsgálva felállítható egy általánosított eukarióta sejtciklus modell, de számos esetben a jellemző eltérések alapján különböző sejtciklus-típusok írhatók le( 2–5. ábrák).
A soksejtű szervezetekben tipikusnak tekinthető sejtciklus négy szakaszra, sorendben G1, S, G2 és M fázisokra tagolható.
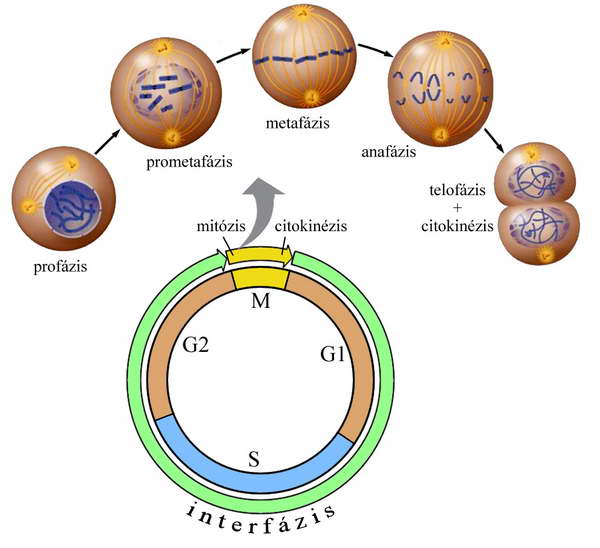
1. ábra Általánosított eukarióta sejtciklus fázisai és az M fázis szakaszai.
Emlős sejtek szövettenyészetében egy ciklus kb. 16–24 órát vesz igénybe. Az S, G2 és M fázisok hossza kevéssé változó, legrövidebb az M fázis, leghosszabb és a körülményektől függően változó a G1.
A sejtciklus legalapvetőbb funkciója a sejt örökítő anyagának megkettőzése, a DNS pontos replikációja, majd ezt követően a kromoszómákba szerveződő DNS kópiák precíz szétosztása két genetikailag egyező utódsejtbe. A DNS szintézise az S fázisban, a kromoszómák szegregációja, a sejtmag- és a sejt kettéosztódása az M fázisban játszódik le.
A legtöbb esetben ahhoz, hogy a sejt számára az osztódáshoz (két életképes utódsejt létrehozásához) szükséges mennyiségű anyag és organellum rendelkezésére álljon, a sejteknek növekedniük kell. Ezért általában az M és S fázis nem követik egymást szorosan, közöttük növekedési fázisok alakulnak ki. Ezek időigényes és jellemző anyagcsere-folyamatokat tartalmazó szakaszok. Az M és S fázis között a G1, az S fázis és a mitózis között a G2 fázis helyezkedik el. Az M fázist megelőző G1, S és G2 alkotja az interfázist, amelyben változó intenzitással, de folyamatosan zajlik a növekedés. Jellemzője az intenzív transzkripciós tevékenység és az erős szintetikus aktivitás.
Az interfázisban több ellenőrző mechanizmus is működik, melyek lehetővé teszik, hogy a sejt monitorozza környezetét és egyben felügyelje a ciklus részfolyamatainak teljesítését.
Egy a mitózisból kilépő, újonnan létrejövő sejt csak akkor lép be egy újabb ciklusba, annak G1 fázisába, ha külső környezete kedvező – pl. megfelelő a tápanyag ellátottság, az oxigénszint stb. –, továbbá jelen vannak (külső eredetű) mitogén hatású szabályozó molekulák. Utóbbiak az állati sejtek esetében az adott sejtre specifikus növekedési faktorok. A mitogén szignált a sejt felszínén kifejeződő receptor molekulák kötik meg és internalizálják. A jelátvezetési útvonal sokszereplős, végül a sejtmagban az ún, korai gének aktiválását eredményezi. A géntermékek elindítják az S fázist előkészítő, valamint a citoplazma növekedését, a sejtorganellumok gyarapodását biztosító folyamatokat.
A sejt citoplazmatikus komponenseinek, organellumainak utódsejtekbe való kerülése nem olyan pontos, mint a genetikai anyagé, ezért az utódsejtek tömege némileg eltérhet. Ez a különbség már a következő ciklus G1 fázisában kiegyenlítődhet úgy, hogy a sejt hosszabb időt tölt a növekedési fázisokban.
A lezajló anyagcsere folyamatok hatására a sejt a G1 fázis végén, de még az S fázis megkezdését megelőzően "elkötelezetté válik" a DNS szintézis megkezdésére. Ez a restrikciós pont (magasabb rendűeknél; élesztőknél Start ellenőrzési pont). E ponton túl már visszavonhatatlanul megindul az S fázis végrehajtásához szükséges fehérjék szintézise, a ciklus eseményei már automatikusan követik egymást, és a ciklus csak akkor áll le, ha a folyamatokba kijavíthatatlan hiba csúszik, vagy a sejtet külső eredetű sérülés éri.
A restrikciós pontot megelőzően az extracelluláris miliő változása vagy a mitogén szignál hiánya következtében a sejt G1-ből kiléphet G0 fázisba. Ez a lépés lehet válasz a sejtet ért kedvezőtlen változásokra, de lehet program szerinti, a differenciálódást előkészítő változások hatása is. A G0 fázisú sejtek szintetikus kapacitását már nem anyagaik megkettőzése, hanem differenciálódásuknak megfelelően, funkciójuk ellátásához szükséges anyagok előállítása köti le. A soksejtű szervezetekben a sejtek többsége G0 fázisban van, ahol napokat, heteket, sőt éveket is eltölthetnek. A differenciált sejtek jelentős része elveszíti osztódó képességét (pl. idegsejtek), egy részük még megfelelő inger(ek) hatására képes rövid ideig visszalépni a ciklusba (pl. májsejtek). Gyorsan cserélődő, megújuló szövetekben (pl. a bélhám vagy a bőr epidermis rétege) a folyamatosan pusztuló differenciált sejtek között vagy alatt differenciálatlan ún. törzssejtek biztosítják a sejtutánpótlást.
A sejtek G0-ból a ciklusba történő visszalépése mindenképpen a restrikciós pont előtt kell megtörténjen. G1-ből S fázisba akkor léphet a sejt, ha növekedési faktorokhoz jutott, megfelelő tömegűre nőtt, a DNS állomány sértetlen, és a replikációhoz szükséges anyagok és apparátus rendelkezésre áll.
Az S fázisban zajlik a DNS replikáció és annak állandó ellenőrzése. A DNS-ről csak egyetlen másolat készülhet, amit molekuláris mechanizmusok biztosítnak és folyamatosan ellenőriznek. Megkettőződik a sejtközpont, benne a centriólum is, de az M fázisig még egymás közelében maradnak.
A G2 fázisban elsősorban a mitózis lebonyolításához szükséges fehérjék előállítása zajlik. A fázis végén található ellenőrzési pont a DNS megkettőződésének hibátlansága, a DNS épsége és az M fázishoz szükséges anyagok rendelkezésre állása esetén engedi továbblépni a sejtet az M fázisba.
Az M fázisban zajlik a mitózis (magosztódás, kariokinézis) és a sejttest befűződéssel történő kettéválása (citokinézis).
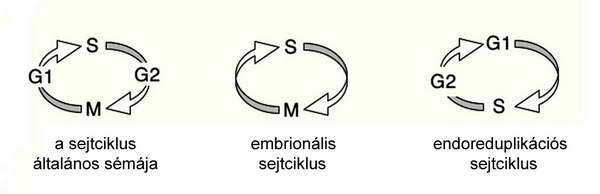
2. ábra A soksejtű szervezetek sejtciklusának néhány jellegzetes típusa.
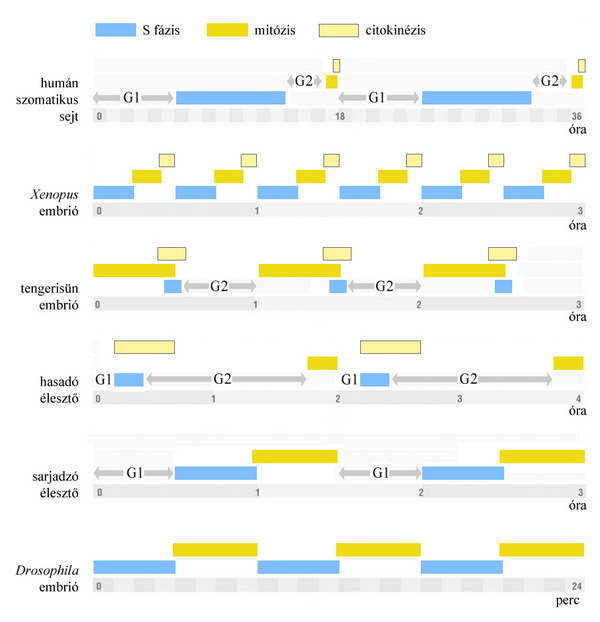
3. ábra Néhány modell szervezet sejtciklusának idődiagramja.
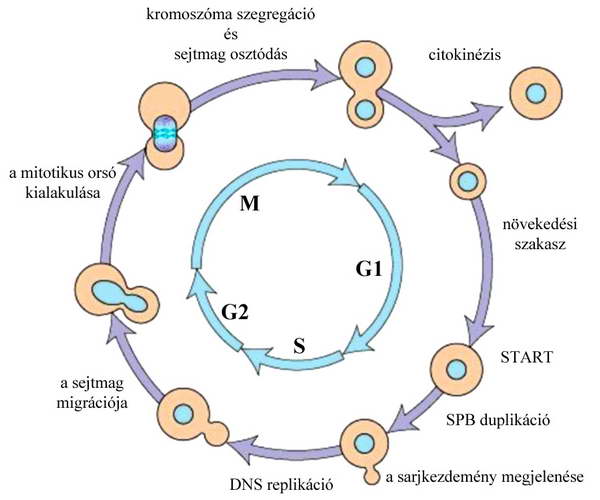
4. ábra A sarjadzó élesztő (Saccharomyces cerevisiae) sejtciklusa fázisai és legjellegzetesebb eseményei.
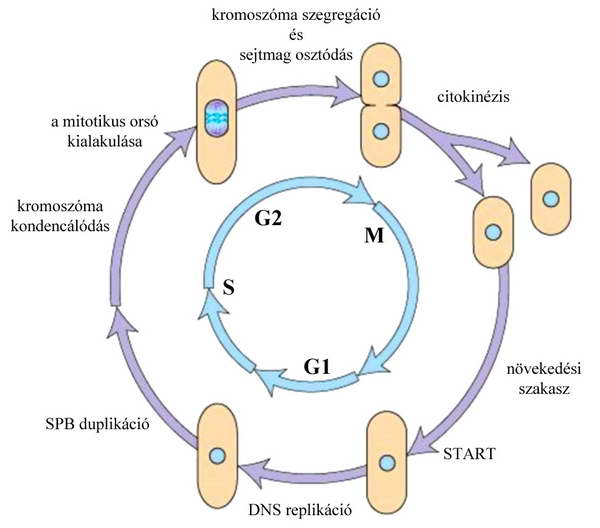
5. ábra A hasadó élesztő (Schizosaccharomyces pombe) sejtciklusának fázisai és jellemző eseményei.