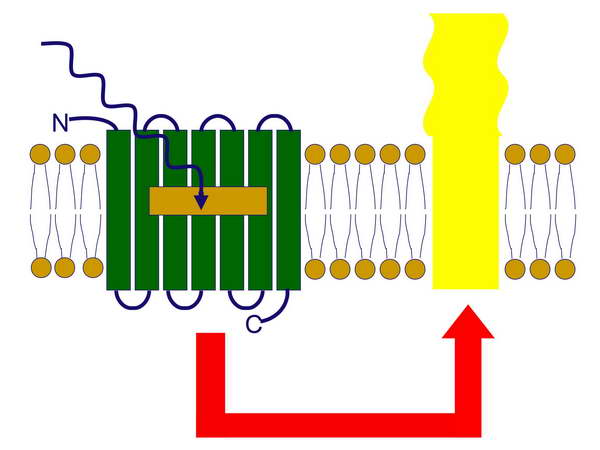
Mint láttuk azt az első sejtről (LUCA) szóló fejezetben, az elektrontranszporthoz, proton-pumpáláshoz és proton elektrokémiai potenciálkülönbséghez kapcsolt ATP-szintézis már jelen lehetett a Föld korai körülményei között. A H+ grádienset a korai sejtek már hasznosíthatták molekulák transzportjára is az ATP szintézise mellett. Az Archeák közé tartozó sótűrő Halobaktériumok rendelkeznek egy olyan fehérjével a membránjukban, amely egy fény-indukált proton-pumpa, a foton energiáját protonok membránon keresztüli pumpálásásra, azaz elektrokémiai potenciálkülönbség generálására használják fel. A membránban elhelyezkedő ATP-szintáz pedig ezt a potenciál-különbséget ATP szintézisben realizálja. A bakteriorodopszin kromofórja az all-trans retinál, amely a foton elnyelését követően ledisszociál a fehérjéről (opszin), amely aktív konformációt vesz fel és proton-pumpaként működik (27-29. ábrák).
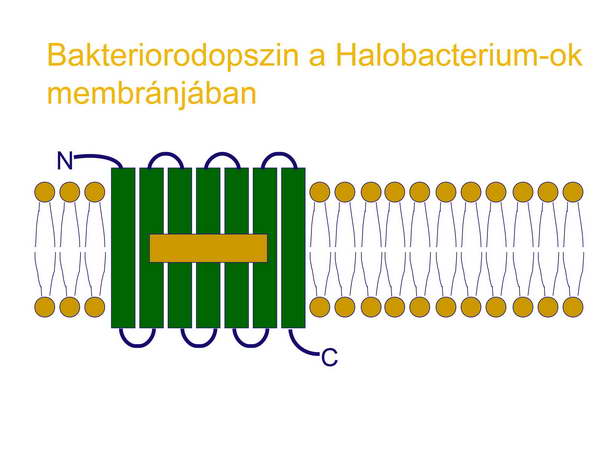
27. ábra
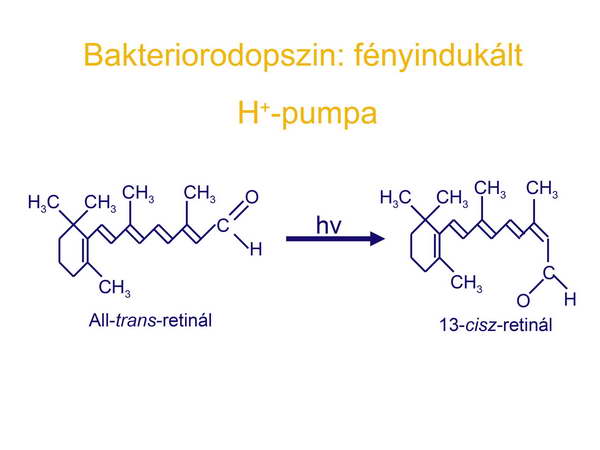
28. ábra

29. ábra
Érdemes megemlíteni, hogy a Halobaktériumok rendelkeznek egy másik típusú fényérzékeny fehérjével (érzékelő rodopszin), amely orientációs szignálként hat a baktériumsejtek számára. Abban az esetben, ha a „kedvező” fény fotonjai érkeznek a kromofórra, a baktérium a fény irányában fog úszni, ellenkező esetben pedig a fényforrással ellentétes irányban. Mindez azért lehetséges, mert ezekben a sejtekben a fény észlelését6 követően egy szignalizációs rendszer indul be, amely a rodopszinból a bakteriális ostor felé továbbítja a szignált. Az úszás irányának megváltoztatását az ostort mozgató rotor forgási irányának megváltozásával éri el a baktérium (30. ábra).
30. ábra