Tartalom
Az eukarióta sejt kialakulásáról szóló fejezetben láttuk, hogy a horizontális géntranszfer következtében mind a mitokondrium, mind a kloroplasztisz génállományának (organelláris genom) jelentős része átkerült a „gazdasejt” genomjába (nukleáris genom), amely a sikeres szimbiózist követően a központi irányító és információ-tároló funkciót egyaránt ellátja. Ennél fogva az organelláris fehérjék jelentős része a nukleáris genomban kódolódik, ott íródik át mRNS-sé, és citoplazmikus riboszómákon transzlálódik fehérjévé. Ezeknek a fehérjéknek be kell jutni az organellumokba, amelyre az evolúció során transzlokációs-mechanizmusok alakultak ki mind a mitokondriumban (Translocator of outer membrane: „Tim” és Translocator of outer membrane „Tom”), mind a kloroplasztiszban (Translocator of outer chloroplast membrane „Toc” és Translocator of inner chloroplast membrane „Tic”). Ezek evolúciósan egymástól függetlenül jöttek létre, de hasonlóan működnek. A két organellumban működő mechanizmusok közös sajátossága, hogy a transzlokálandó fehérje egy szignál-szekvenciával együtt érkezik az organellum külső membránjához egy citoplazmikus riboszómáról, és ezt a szignál-szekvenciát egy speciális felismerő fehérje felismeri. Ezt követően a külső membrán transzlokációs komplexe addig diffundál a külső membrán síkjában, amíg egy síkba nem kerül a belső membrán transzlokációs komplexével, majd chaperonok és ATP közvetítésével a fehérje egyszerre hatol át mindkét membránon (1. ábra).
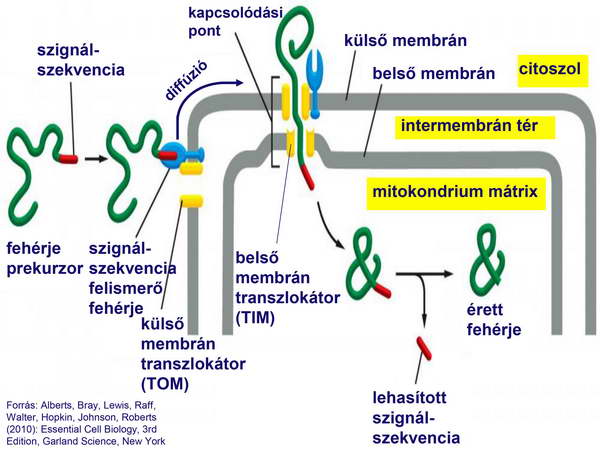
1. ábra
A sejten belüli üzenetküldés (szignalizáció) iránya tehát normális körülmények között a sejmagtól a periféria felé (a nukleáris genomból az organellumok felé) mutat. Ezt a típusú üzenetküldést anterográd szignalizációnak nevezzük, és legfontosabb feladata a saját genommal rendelkező organellumok működőképességének biztosítása és fenntartása. Az egyes organelláris fehérjék többször több alegységből épülnek fel, és előfordul, hogy ezek az összetett fehérjék genetikai „kimérák”, azaz különböző alegységeik különböző genomikus eredetűek. Ennek egyik legnevezetesebb példája a (C3-as növényekben a) kloroplasztisz egyik legfontosabb enzimének, a ribulóz-bifoszfát-karboxiláznak (RUBISCO) az esete. Ennek a nagyobbik alegysége (Large Subunit, LS) a kloroplasztisz genomban kódolódik és kloroplasztikus riboszómákon transzlálódik, a kisebb alegység (Small Subunit, SS) viszont a nukleáris genomban kódolódik és citoplazmikus riboszómákon transzlálódik. Mindez lehetőséget ad a két komponens arányának mindenkori optimális szabályozására. Ha ugyanis a nagyobb alegységből termelődik több, akkor a „felesleg” a kloroplasztiszon belül represszálja a további fehérjeszintézist (negatív „feedback”). Ha viszont a nukleáris genomban kódolt kisebb alegységből termelődik több, akkor ez transzlációs aktivátorként indukálni tudja az organelláris kódolású nagyobb alegység expresszióját
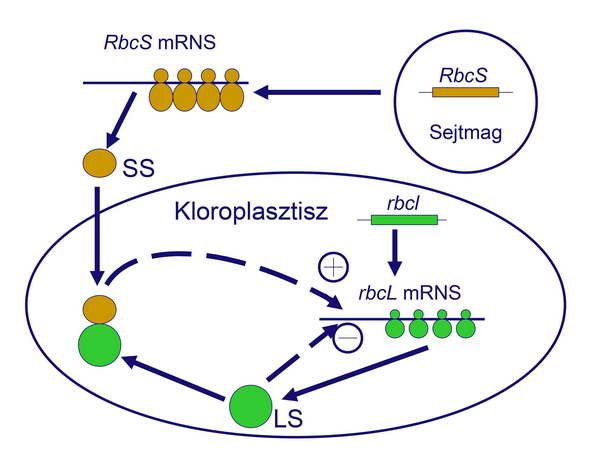
2. ábra
Az anterográd szignalizáció szép pédája: a RUBISCO alegységeinek összeállása.
A transzkripciós faktorok és az enhancer szekvencia működése
Az 1 receptor - 3 szignalizációs útvonal szemléltetése
A MAP-kináz szignalizációs kaszkád működése
Az enhancer szekvencia közelébe kerülő gén up-regulációja reciprok kromoszóma-transzlokációt követően