Tartalom
- A biológiai membránok elválasztó funkciója
- A biológiai membránok szerveződésének alapjai
- A biológiai membránok dinamikai sajátságai
- A biológiai membránok áteresztőképessége (permeabilitása)
- A biológiai membránok elektromos tulajdonságai
- A membrán két oldala közötti egyenlőtlen ioneloszlás.
- A membránnak az egyes ionokkal szembeni eltérő permeabilitása
- A membránnak az intracelluláris közegben levő fehérjemolekulákkal szembeni impermeabilitása
- A membránon keresztüli elektrogén (elektromosságot termelő) ion-transzport folyamatok.
- A biológiai membránok összekötő funkciója
- A membránfolyamatok sajátosságai növénysejtekben
A biológiai membránok minden élő sejt különlegesen fontos képződményei, amelyekről túlzás nélkül állítható, hogy „két világ határán” foglalnak helyet. A két világot mindig két olyan közeg jelenti, amelyek eltérő összetételűek, néha szinte alig van olyan paraméter, amiben hasonlítanának egymásra. E két különböző közeg határán a membránok alapvető feladata, hogy mindkét közeg sajátos összetételét megőrizzék és fenntartsák a legkülönbözőbb környezeti viszonyok között is. Bár a növénysejteknek van sejtfaluk, a tulajdonképpeni határfelületet a citoplazma és a külvilág között a plazmalemma képezi. A plazmalemma egyfajta speciális biológiai membrán, ezért a plazmalemma specifikumainak tárgyalása előtt áttekintjük a biológiai membránok szerveződésére vonatkozó alapvető ismereteket.
A biológiai membránok alapfunkciója, hogy két víztartalmú közeget úgy különítsenek el egymástól, hogy bizonyos anyagok számára biztosítsák az átjutást egyik közegből a másikba. A membránok tehát paradox módon egyszerre elkülönítenek, és össze is kötnek két közeget. Minderre speciális felépítésük teszi képessé őket. A biológiai membránok alapvázát egy lipid kettősréteg képezi, amelyben diffúz módon fehérjemolekulák helyezkednek el. A membránok alkotórészei között alapvető jelentőségűek a lipidtermészetű anyagok, közülük is az ún. poláris lipidek. Ezek poláros és apoláros csoportokat egyaránt tartalmazó, amfifil karakterű anyagok. Poláris fejcsoportjaik révén a vizes fázissal képesek asszociálódni és elektrosztatikus kölcsönhatásokat kialakítani, míg az apoláros zsírsav-oldalláncok között fellépő Van der Waals- és hidrofób kölcsönhatások összetartó erőt képviselnek és hozzájárulnak a membrán belső, apoláros régiójának stabilizálásához.
A lipidmolekulák asszociációja nem csak a biológiai membránok felépítésében nyilvánul meg. A poláris lipidmolekulák két-dimenziós struktúrákat létrehoznak vizes fázis felszínén is, ezek orientációjuk alapján egyrétegű filmek vagy köralakú micellák lehetnek. A membrán-kettősréteg gyűrűvé záródásával alakul ki a 3-dimenziós liposzóma, melyek belsejében egy vizes fázist tartalmazó üreg van. A liposzómát határoló lipid kettősréteg a sejtmembránhoz hasonló felépítésű, ezért a liposzómákat frekventáltan hasznosítják a gyógyászatban különféle nagymolekulájú gyógyszerek sejtbe történő bejuttatására, vagy a kozmetikai iparban különféle bőrtápláló anyagoknak a bőrhámsejtekbe történő bevitelének megkönnyítésére.
A membránalkotó lipidek lényegi sajátossága, hogy aktuálisan milyen fejcsoportok és milyen zsírsavláncok találhatók az illető lipidféleségben. A fejcsoportok alapján az egyik legfontosabb lipidosztály a foszfolipidek, melyekben a glicerin szabad OH-csoportját foszforsavmaradékhoz kapcsolódva észteresíti a fejcsoport (foszfatidil-kolin, foszfatidil-etanolamin, stb.). Az állati sejtek membránjaiban döntően foszfolipidek fordulnak elő, és ugyancsak foszfolipidek dominálnak a növénysejtek plazmalemmájában és endoplazmatikus retikulum membránjában is. Lényeges különbség viszont, hogy a kloroplasztisz belső membránjaiban (tilakoidok) galaktolipidek fordulnak elő döntő mennyiségben. Ezekben a glicerin szabad OH-csoportjához galaktóz-molekulák kapcsolódnak. Azt a lipidet, amelyhez 1 galaktóz-molekula csatlakozik, monogalaktozil-digliceridnek (MGDG), míg a 2 galaktóz-molekulát tartalmazó lipidet digalaktozil-digliceridnek (DGDG) nevezzük. E két lipidféleségnek igen sajátos a zsírsavösszetétele is: dominál bennük a 2 kettőskötést tartalmazó linolsav (18 szénatom:2 kettőskötés = 18:2) és a 3 kettőskötést tartalmazó linolénsav (18:3). Mindkét zsírsav alapvető fontosságú az állati szervezetek számára is, hiszen az állati sejtekben a hosszú szénláncú és több kettőskötést tartalmazó zsírsavak (pl. arachidonsav (20:4), eikoza-pentaénsav (20:5), dokoza-hexaénsav (22:6)) bioszintéziséhez linolsavra, illetve linolénsavra van szükség, ezek az állati sejtek számára eszenciális zsírsavak.
A biológiai membránok másik alapkomponensei a membránfehérjék, melyek között a membránon belüli topológiai elhelyezkedés alapján megkülönböztetünk periferiális (a membrán poláros fejcsoport-régiójával asszociálódó), illetve integráns, azaz a membrán apoláris régiójába is belenyúló fehérjéket. Amennyiben egy fehérje a membrán teljes síkját átíveli, transzmembrán fehérjéről beszélünk. A fehérjék közvetlen környezetében elhelyezkedő lipidréteg (annuláris lipid) milyensége (fejcsoportja és zsírsav-összetétele) döntő az illető fehérje funkcióképessége szempontjából. A membránfehérjék funkciójuk alapján lehetnek receptorok, transzport-fehérjék, enzimek (pl. kinázok, foszfatázok), valamint a citoszkeleton elemei (pl. aktin-filamentumok).
A biológiai membránok dinamikai sajátságai az egyes komponensek mozgásával és mozgékonyságával kapcsolatos jelenségeket tükrözik. Közismert, hogy az anyagok állandó mozgásban vannak és ez érvényes a membránt felépítő lipid- és fehérjemolekulákra is. A membránlipidek alapvetően háromféle mozgástípust végezhetnek a membránban, melyek a rotáció, azaz az egész molekula saját tengely körüli forgómozgása, a laterális diffúzió, azaz a molekuláknak a membrán kétdimenziós síkjában történő elmozdulása, valamint a flip-flop, ami egy lipidmolekulának az egyik membránfélből a másikba történő átkerülését jelenti. A membránban levő fehérjemolekulák ugyancsak végeznek rotációs forgómozgást és laterális diffúziót, azonban a membránfehérjék átkerülése egyik membránfélből a másikba meglehetősen ritka és valószínűtlen esemény. Megjegyzendő, hogy a membránfehérjék laterális diffúzióval jutnak el a membrán egyik helyéről egy másikra, és tulajdonképpen ez képezi az alapját a fehérje-fehérje interakcióknak, amelyek pl. a receptor-G protein kölcsönhatás során megvalósulnak.
Azt, hogy az egyes komponensek mennyire szabadon mozdulhatnak el egy adott membránon belül, a membrán fluiditása, illetve rigiditása határozza meg. Minél fluidabb egy membrán, annál könnyebben mozdulnak el benne az egyes komponensek. Maga a fluiditás és a rigiditás a membrán fizikai-kémiai állapotával függ össze. A membrán egy komplex rendszer, amelynek halmazállapota egy folyadék-kristályhoz hasonlítható. Benne talalhatunk olyan részeket („szigeteket”), amelyek folyékonyabbak (fluidabbak) és olyanokat, amelyek szilárdabb (rigidebb vagy inkább gél-szerű) halmazállapotúak. A membránnak ezt a tulajdonságát egyrészről a lipidek és főleg a zsírsavak, másrészről pedig az aktuális hőmérséklet határozza meg. Alapszabály, hogy minél több a telített zsírsav az adott membránban vagy membránszakaszon, annál rigidebb az adott membrán. Ez fordítva is igaz: a telítetlen zsírsavak jelenléte növeli a membrán fluiditását. Mindezt egyszerűen értelmezhetjük, ha elképzeljük egy adott lipidmolekula rotációs forgómozgását. A telített zsírsavláncok a szénatomok közötti egyes kötés miatt, bár zegzugos, de egyenes lefutásúak, azaz a saját tengelye körül forgó lipidmolekula hengerszimmetrikus. Ez a szerkezeti sajátosság jellemző a foszfolipidekre és lehetővé teszi a foszfolipidek számára, hogy kettősrétegű membránstruktúrákat alakítsanak ki. A telítetlen zsírsavláncokban levő kettőskötés azonban, amennyiben cisz konfigurációjú, a C-C kötések lefutását megváltoztatja és a kettőskötés után a zsírsavlánc megtörését okozza. Egy ilyen lipidmolekula rotációs forgómozgásához sokkal nagyobb sugarú térrészre van szükség, az ilyen lipidmolekula nem henger-, hanem kúp-szimmetrikus. Az ilyen lipideket tartalmazó membránokban a területegységre jutó lipidmolekulák száma kevesebb, azok „lazábban pakolva” helyezkednek el a membránban. Ez a lazább szerkezet értelemszerűen megkönnyíti a szomszédos lipid- és fehérjemolekulák elmozdulását is, végeredményben tehát abban a membránrészben, ahol kettőskötést is tartalmazó zsírsavakat hordozó lipidek fordulnak elő, nagyobb lesz a membránalkotók mozgási szabadsága, azaz fluidabb lesz az adott membrán. Könnyen belátható, hogy már egyetlen kettőskötés jelenléte is jelentősen deformálja a lipidmolekula hengerszimmetriáját, az ilyen lipidek forgási terét leginkább egy kúppal lehetne jellemezni, melynek csúcsában helyezkedik el a lipidmolekula fejcsoportja, palástját pedig a forgó zsírsavláncok felülete alkotja. Ez a szerkezeti sajátság leginkább a galaktolipidekre (MGDG és DGDG) jellemző, hiszen bennük dominálnak a 2, illetve 3 kettőskötést tartalmazó zsírsavak, a linol-, illetve a linolénsav. Ezek a lipidek szinte kizárólag a kloroplasztisz belső membránjaiban, a tilakoidokban találhatók. Mivel ezek a kúpalakú lipidmolekulák egymás mellé pakolva hajlott, görbült membránfelszíneket hoznak létre, kézenfekvőnek tűnik az a feltételezés, hogy alapvető szerepük van a tilakoidmembránok görbületeinek kialakításában. A kloroplasztisz-membránok lipid- és zsírsavmintázata tehát egyrészt a membránok extrém magas fluiditását, másrészt a görbült membránfelszínek kialakítását is lehetővé teszi.
Említettük, hogy a membrán dinamikai sajátságainak másik alapvető meghatározója az aktuális hőmérséklet. Ennek az az oka, hogy a hőmérséklet közvetlenül meg tudja változtatni a membránok fizikai-kémiai állapotát. Minden membrán jellemezhető egy hőmérséklet-értékkel, amely környezetében a folyadék-kristályos fázis egyensúlyt tart a szilárd fázissal. Ennél alacsonyabb hőmérsékleten a szilárd fázis, míg ennél magasabb hőmérsékleten a folyadék-kristályos fázis dominál. Ezt a hőmérséklet-értéket az adott összetételű membrán vagy membránrész tranzíciós (fázisátmeneti) hőmérsékletének nevezzük. Ez a paraméter a membránműködés szempontjából döntő fontosságú, hiszen a membránban levő fehérjemolekulák csak addig képesek laterális diffúzióra, amíg a membrán folyadék-kristályos, azaz félfolyékony halmazállapotú. A rigid, merev, gélszerű halmazállapotú membránban a komponensek mozgási szabadsága minimálisra csökken, a membrán „megdermed”, ezért ez az állapot a membrán működésével összeegyeztethetetlen. Egy adott membrán tranzíciós hőmérséklete alapvetően attól függ, hogy az adott membránrészben levő lipidekben mennyi a telítetlen zsírsavak aránya a telített zsírsavakhoz viszonyítva. Általános szabály, hogy minél több a telítetlen zsírsav egy adott membránszakaszon, annál alacsonyabb annak a membránszakasznak a tranzíciós hőmérséklete, azaz annál alacsonyabb hőmérsékleten lesz a folyadék-kristályos fázisból szilárd, gél halmazállapotú fázis, tehát annál alacsonyabb hőmérsékleten dermed meg a membrán. Ez más szóval azt jelenti, hogy minél telítetlenebb egy membrán, annál alacsonyabb hőmérsékleten marad meg a folyadék-kristályos szerkezete, ami az életfolyamatok szempontjából alapvető membránműködés alapfeltétele.
A sejtek számára a környező közeg hőmérséklete elsősorban a membránszerkezet modulációja és az ezzel kapcsolatos funkcionális változások miatt fontos. Az állandó testhőmérsékletű szervezetekben a membránok lipid- és zsírsavösszetétele az állandó testhőmérséklet értékének megfelelően alakult ki és az adott hőmérsékletnek megfelelő szinten optimalizálódott. A növények közismerten nem képesek testhőmérsékletüket szabályozni, ezért a növénysejtek membránjai fokozottan érzékenyek a környező hőmérséklet hirtelen megváltozására. Különösen a hideg- és fagyhatás az, ami a növénysejteket fokozottan próbára teszi. Mint láttuk, a hőmérséklet csökkenése a membrán fázisállapotának megváltoztatásával (a fluiditás csökkentésével) fejti ki sejtkárosító hatását, és a növényfajok többsége a hirtelen lehűlést és fagyot nem is tudja elviselni, bennük visszafordíthatatlan működésbeli zavarok lépnek fel, amelyek a sejtek funkciókiesését, később pedig pusztulását okozzák. Van azonban néhány olyan faj és fajon belüli változat, amelyek bizonyos mértékig ellenállóak a hirtelen hideg- vagy fagyhatással szemben. Ezeket a növényeket nevezik hidegtűrő, illetve fagyálló növényeknek. Bebizonyosodott, hogy ezek a növények hőmérsékleti adaptációjuk során olyan mechanizmusokat hoznak működésbe, amelyekkel csökkenteni tudják membránjaik tranzíciós hőmérsékletét. Egyrészt ezek a fajták normál körülmények között is olyan zsírsavösszetétellel rendelkeznek, amelyben több a telítetlen zsírsav, ílymódon ezek a membránok már „felkészülten” várják az esetleges hirtelen hőmérséklet-csökkenést. Ezen túlmenően, a hirtelen hideghatás egy szignálként hat ezekben a növényekben, melynek hatására olyan génszakaszok íródnak át, amelyek termékei speciális enzimek. Ezek az enzimek kettőskötés kialakítására képesek a telített zsírsavláncokban, összefoglaló néven deszaturázoknak nevezik őket. A génexpresszió révén megjelenő deszaturázok azután a plazmalemmához és a többi intracelluláris membránrégióhoz szállítódnak és ott további kettőskötéseket alakítanak ki, ezzel jelentősen hozzájárulva a membránok tranzíciós hőmérsékletének csökkentéséhez.
A növényi membránok (különösen a kloroplasztisz tilakoidmembránjai) sajátos lipid- és zsírsavösszetétellel rendelkeznek, melynek fontos humán táplálkozásélettani szerepe van. 1/ A többszörösen telítetlen növényi zsírsavak eszenciális zsírsavak számunkra. 2/ Ha étrendünkben növeljük a többszörösen telítetlen zsírsavak részarányát, ezek beépülve membránjainka, fluidabb membránokat hoznak létre, amelyek számos kóros elváltozás (szív- és érrendszeri betegségek) esetében igen fontos preventív szerepet töltenek be. Ezen a helyen kell kiemelnünk a növényi membránok (elsősorban a kloroplasztisz-membránok) azon sajátságát, hogy gyakorlatilag nem tartalmaznak koleszterint.
Közismert, hogy a koleszterin feldúsulása membránjainkban nem kívánatos, hiszen súlyos megbetegedések (szívinfarktus, érelmeszesedés) egyik kockázati tényezőjéről van szó. Mindez képezi az ún. „reform-táplálkozás”, azaz a koleszterinmentes, többszörösen telítetlen zsírsavakban dús növényi táplálék fogyasztásának táplákozásélettani alapját. Sajnos, azonban, a többszörösen telítetlen zsírsavak nemcsak áldást hoznak számunkra, hanem igen komoly veszélyforrást is jelentenek. Mindez az ún. oxidatív stressz mechanizmusával kapcsolatos. A sejtekben az oxidatív folyamatok során mindig termelődnek ún. oxigén szabad gyökök. Ezek párosítatlan elektronnal rendelkező képződmények, tehát olyanok, amelyek elektronszerkezetében van egy olyan elektron, amelynek nincs meg az ellentétes spin-kvantumszámú párja. Ezek a szabad gyökök éppen azért annyira reakcióképesek, mert keresik az ellentétes spin-kvantumszámú párjukat. A sejtekben elsősorban a mitokondriumok belső membránjában keletkeznek, amikor is az oxidatív reakciólánc végén a molekuláris oxigén redukciójára (elektronnal történő találkozására) kerül sor. A folyamat eredménye víz, azonban bizonyos valószínűséggel a molekuláris oxigén és az elektron reakciójából szuperoxid szabad gyökök is képződnek. Ezek már önmagukban is igen reakcióképes oxidálószerek, azonban sokkal veszélyesebb tulajdonságuk, hogy további szabad gyökök (pl. hidroxil anion gyök) termelődését indítják be, amelyek még agresszívebb oxidálószerek. Összefoglaló néven az oxigénből keletkező szabad gyököket reaktív oxigén fajtáknak (ROS) nevezzük. Ezek előszeretettel támadják meg a zsírsavláncok telítetlen kettőskötéseit, azokat oxidálják és peroxidálják, ezáltal egyrészt megváltoztatják a membrán zsírsavösszetételét és ezzel annak dinamikai sajátságait, másrészt a lipidperoxidáció révén visszafordíthatatlan folyamatot indítanak el, amelynek eredményeként tartós, maradandó membránkárosodás következik be. Mivel a membránszerkezet integritása a belső környezet fenntartásának egyik legalapvetőbb feltétele, mindenfajta membránkárosodás a sejt számára letális veszélyforrást jelent és előbb-utóbb a sejt halálához vezet.
A membrán permeabilitásán vagy áteresztőképességén az egyes anyagok membránon történő áthaladásának a sebességét értjük. Közismert, hogy vannak olyan anyagok, amelyek a membránon keresztül viszonylag akadálytalanul hatolnak át, de vannak olyanok, amelyek számára ez az áthatolás kisebb-nagyobb mértékben nehézségekbe ütközik. Az ilyen anyag számára a membrán jelentős akadályt jelent. Elvileg (végtelen idők alatt) minden anyag átjut minden membránon, azonban az átjutás sebessége néha végtelenül lassú. Szakkifejezéssel élve ezekre az anyagokra a membrán nagyon kevéssé vagy egyáltalán nem permeábilis, azaz impermeábilis. A vizet a biológiai membránok sokkal könnyebben átengedik, mint a benne oldott anyagokat, különösen, ha azok mérete igen számottevő (pl. fehérjemolekulák). Ezt a sajátságot fejezi ki a membrán félig áteresztő (szemipermeábilis) jellege.
A membránon történő átjutás tulajdonképpen kettős akadály legyőzését jelenti. A membrán két vizes közeg határán levő alapvetően hidrofób karakterű elválasztó réteg, mely éppen a hidrofób karaktere miatt képes két vizes közeg elkülönítésére és az elválasztó funkció betöltésére. Ezen túlmenően azonban a membrán felszíne elektromos töltést hordozó lipid-fejcsoportokból áll, amelyeket egy elektromosan szigetelő hidrofób réteg választ el egymástól. Ez az elrendezés teljesen analóg az elektronikából ismert kondenzátorral. A membrán tehát a két vizes fázis között elektromos szigetelőként viselkedik. Ez az elektromos szigetelőréteg nem befolyásolja az elektromosan semleges molekulák (pl. cukrok) átjutását, de jelentősen megnehezíti az elektromosan töltött részecskék, mindenekelőtt az ionok keresztüljutását a membránon.
Ami a membránt mint apoláros elválasztó-közeget illeti, általános szabály, hogy rajta könnyen áthatolnak az apoláros, lipidoldékony anyagok (pl. szerves oldószerek és gázok), viszont a poláros, hidrofil anyagok számára - különösen, ha azok mérete számottevő - a membránok igen nehezen átjárhatók. Ezek közé tartozik az összes vízben oldott anyag, tehát cukrok, aminosavak, de mindenekelőtt a vízben oldott sók disszociációjából, illetve hidrolíziséből származó ionok. Úgy tűnik tehát, mintha a membrán „szelektálna” és előnyben részesítené a hidrofób anyagokat. Ez mindenekelőtt a membrán lipidfázisának hidrofób karakterével magyarázható és jól felhasználható annak eldöntésére, hogy egy adott anyag várhatóan milyen lassan vagy gyorsan halad át a biológiai membránokon. Ez a kérdés gyakran felvetődik pl. a gyógyszeriparban, amikor egy készítményről tudni kell, hogy milyen sebességgel halad át a membránokon. Ennek becslésére szolgál az olaj/víz megoszlási hányados meghatározása. Az illető anyagot feloldják olajban is és vízben is, és meghatározzák az oldatba ment (feloldódott) anyag koncentrációját mindkét oldószerben. E két érték hányadosa adja meg az olaj/víz megoszlási hányadost. Kimutatták, hogy egyenes arányosság áll fenn egy adott anyag olaj/víz megoszlási hányadosának értéke és egy adott biológiai membránon történő áthaladásának sebessége között. Minél nagyobb az olaj/víz megoszlási hányados értéke, annál gyorsabban képes az illető anyag áthatolni a membránon, azaz annál gyorsabban kerülhet az extracelluláris közegből a sejt belsejébe.
Különleges sajátsága a biológiai membránoknak, hogy a víz könnyebben halad át rajtuk, mint az a termodinamikai megfontolások alapján elvárható lenne. Ez a megfigyelés vezetett ahhoz a feltételezéshez, hogy a membránban kell, hogy legyenek speciális csatornák, amik megkönnyítik a vízmolekulák átjutását. A legutóbbi kutatások valóban megerősítették, hogy ilyen csatornák valóban léteznek. Aquaporinoknak nevezik őket.
Ha az élő sejt membránjain is passzív diffúzióval jutnának át az anyagok, az sok esetben lehetetlenné tenné a normális sejtműködést, és ez lenne a helyzet, hogyha a biológiai membránok pusztán poláris lipidekből álló kettősrétegek lennének. „Szerencsére” a biológiai membránok a lipid kettősrétegbe ágyazódva fehérjéket is tartalmaznak, és a membránfehérjék jelentős részének éppen az a feladata, hogy a passzív diffúzióval nehezen, illetve lassan áthaladó anyagok átjutását meggyorsítsák. Ezeket hívjuk összefoglaló néven transzportfehérjéknek, funkcionális tulajdonságaikról a Membrán-transzport fejezetben teszünk említést.
Az előző részben már utaltunk arra, hogy a membrán a lipid-fejcsoportok töltése és a két fejcsoport-réteg között húzódó hidrofób réteg miatt egy elektromos kondenzátorhoz hasonlítható. A fejcsoportok töltéséből adódóan a membrán felszínének közvetlen közelében az ionok az elektrosztatikus kölcsönhatások szerint rendeződnek és egy kettősréteget hoznak létre. Ezt az elrendezést, amelyet elsősorban a lipidfejcsoportok negatív töltése hoz létre, felületi potenciálnak nevezzük. Értelemszerűen a felületi portenciál a membrán mindkét oldalán kialakul és aktuális értéke a felületegységre jutó negatív töltések számától, azaz a lokális töltéssűrűségtől függ.
A membránpotenciál a membrán által elválasztott két vizes oldat között mérhető feszültség (potenciál-különbség). A fizikában feszültségnek nevezik az ellentétes töltések szétválasztásából származó erőt, és a membránpotenciál értelmezésekor is célszerű ezzel kezdeni gondolatmenetünket. Első megközelítésben a membránpotenciál a membrán két oldalán egyenlőtlenül jelenlevő pozitív, illetve negatív töltések viszonyát jelenti, azaz azt, hogy az egyik oldalon több negatív, a másikon több pozitív töltés található. Értelemszerűen az ellentétes töltések elektrosztatikusan vonzzák („semlegesíteni akarják”) egymást, de a közöttük levő membrán ezt fizikailag megakadályozza. Élő sejtekben lényeges, hogy normális körülmények között mindig a sejt belsejében dominálnak a negatív töltések, így az a negatív pólus (katód), az extracelluláris közeg pedig a pozitív (anód), azaz a sejt belseje általában negatívabb a külvilágnál. Ennek igen fontos következménye, hogy az extracelluláris térben lévő kationokra elektrosztatikus vonzóerő, míg az anionokra elektrosztatikus taszítóerő hat. A membránpotenciál kialakításában 4 fő tényező játszik szerepet: 1/ a membrán két oldala közötti egyenlőtlen ioneloszlás; 2/ a membránnak az egyes ionokkal szembeni eltérő permeabilitása; 3/ a membránnak az intracelluláris közegben levő fehérjemolekulákkal szembeni impermeabilitása, és 4/ a membránon keresztüli elektrogén (elektromosságot termelő) ion-transzport folyamatok. A következőkben röviden mind a négy tényezőt megvizsgáljuk.
Az intracelluláris és extracelluláris közeg ionösszetételében mutatkozó jelentős különbség az élő sejtek egyik legfontosabb tulajdonsága, ezen különbség fenntartása pedig az élő sejtek egyik legalapvetőbb funkciója. Az intracelluláris környezet (citoplazma) ionösszetételére magas K+-koncentráció (130-160 mM) és szembetűnően alacsony Na+-koncentráció (5-20 mM) a jellemző, míg a Cl--koncentráció e két szélsőséges érték közötti köztes értéket (30-60 mM) mutat. Az extracelluláris közeg ionösszetétele ettől minden esetben lényegesen eltérő. Az állati sejtek extracelluláris környezetére jellemző, hogy itt a K+ ionok koncentrációja általában igen alacsony (5-10 mM), szemben az igen magas Na+- (130-150 mM) és Cl-- (140-160 mM) koncentrációval. Vizsgáljuk meg most ezek közül a K+ és Cl- ionokra vonatkozó viszonyokat. Képzeljük el, hogy van két edényünk, az egyikben 140 mM-os, a másikban 5 mM-os KCl oldat van, és tételezzük fel, hogy a KCl teljesen disszociál, tehát az egyik oldatban 140 mM a K+ ionok és ugyanannyi a Cl- ionok koncentrációja, míg a másik oldatban mindkét ion koncentrációja 5 mM. Amíg a két edény között semmiféle kapcsolat nincs, addig mindkét edényben az oldatok egyensúlyban vannak, hiszen mindegyikben pontosan annyi pozitív töltésű ion lesz jelen, mint negatív töltésű. Ha azonban a két edény tartalmát egy közös edénybe töltjük úgy, hogy a két edényből származó oldatrészek közé egy féligáteresztő membránt teszünk (pl. celofán-hártyát, ami csak a vizet engedi át, az ionokat egyik irányban sem), a két oldat a membránon keresztül bár, de érintkezik egymással és a kémiai potenciálok (koncentrációk) különbsége miatt azok kiegyenlítésére törekszik. Ha nem lenne jelen a membrán, ez a kiegyenlítődés meg is történne és a koncentrációk pillanatszerű gyorsasággal kiegyenlítődnének. A membrán jelenléte azonban az ionok számára lekűzdhetetlen akadályt jelent, így a koncentráció-különbség diffúzióval történő kiegyenlítése lehetetlenné válik. A két oldat között azonban továbbra is megmarad a „szándék” a koncentráció-különbség kiegyenlítésére, amit egyértelműen bizonyít az, hogy a két oldatfél közötti koncentráció-különbség kiegyenlítődik, ha a két közeget egy elektromos vezetővel kötjük össze. Ekkor a vezetőben áram folyik és az áramot létrehozó feszültség nagysága (a „kiegyenlítődési szándék” erőssége) arányos a fennálló koncentráció-különbség mértékével. Ez a feszültség könnyen detektálható, ha a membrán által elválasztott két térrészbe 1-1 elektródát helyezünk és azokat feszültségmérővel kapcsoljuk össze. A koncentráció-különbségből eredő feszültséget Nernst-potenciálnak nevezzük, melynek értéke a Nernst-egyenlet felhasználásával könnyen kiszámítható: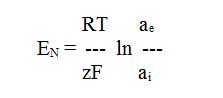
ahol EN a Nernst-potenciál értéke V-ban, R az egyetemes gázállandó, T az abszolút hőmérséklet, z az ion előjeles töltése, F a Faraday-állandó, ae és ai az ion aktivitása (koncentrációval arányos mennyiség) a membránnal elválasztott külső (extracelluláris), illetve belső (intracelluláris) térben. A Nernst-potenciál névlegesen azzal a feszültséggel egyenlő, amely adott koncentrációviszonyok hatására kialakul. Az egyes ionok intracelluláris, illetve extracelluláris koncentrációjának ismeretében a Nernst-potenciál minden egyes ionra külön-külön kiszámítható.
Növénysejtek esetén a helyzet annyiban módosul, hogy a növénysejtek extracelluláris közegére nem jellemző az a stabil összetétel és magas ozmózisnyomás, ami az állati sejtek extracelluláris közegét jellemzi. Általánosságban elmondható, hogy a növénysejtek extracelluláris közege híg oldat, melynek ozmotikus koncentrációja, és így ozmózisnyomása jóval alacsonyabb az állati sejtek extracelluláris közegének megfelelő értékeinél. Mindez elsősorban a NaCl alacsony extracelluláris koncentrációjával magyarázható.
A membránpotenciál másik fontos származási területe a membránnak az egyes ionokkal szembeni eltérő permeabilitása. A biológiai membránok jelentős különbségeket mutatnak az egyes ionokkal szembeni permeabilitásuk vonatkozásában. A biológiai membránok leginkább a K+ ionokkal szemben permeábilisak, legkevésbé pedig a Na+ ionokkal szemben. Ez azt jelenti, hogy a K+-ra fennálló jelentős koncentráció-különbség miatt a K+ ionok sejtből kifelé történő diffúziója igen számottevő, és ez egy diffúziós áramot, valamint egy annak megfelelő diffúziós potenciált hoz létre. Általános szabálynak tekinthetjük, hogy a K+-ionok diffúziós potenciálja az élő sejtek membránpotenciáljának legdöntőbb, meghatározó tényezője. A másik két makroion hozzájárulása a permeabilitási tag alacsony volta miatt nem annyira jelentős. Századunk 40-es éveiben három brit kutató, Goldman, Hodgkin és Katz állt elő azzal az elképzeléssel, hogy állati sejtek, különösen idegdejtek esetében a membránpotenciál a három makroion (K+, Na+ és Cl-) Nernst- és diffúziós potenciáljainak egyfajta kombinációja lehet. Az általuk felállított egyenletben (GHK-egyenlet) az egyes ionok Nernst-potenciáljaihoz hozzárendelték a P-vel jelölt diffúziós tagokat (az adott ionra vonatkozó permeabilitási állandó) és megalkották a Nernst-féle ekvilibrium és a diffúziós potenciálok lineáris kombinációját, mely a kísérleti adatokkal igen jó egyezést mutatott.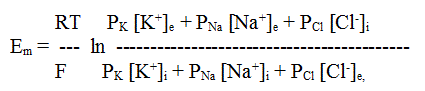 ahol Em a membránpotenciál értéke V-ban, R az egyetemes gázállandó, T az abszolút hőmérséklet, F a Faraday-állandó, [ ]e és [ ]i az adott ion koncentrációja a membránnal elválasztott külső (extracelluláris), illetve belső (intracelluláris) térben, P a membránnak az adott ionra vonatkozó permeabilitási állandója. A membránpotenciál aktuális értéke és az adott ionra vonatkozó Nernst-potenciál közötti mindenkori eltérés fontos információt ad arra vonatkozóan, hogy az adott ion mennyire van egyensúlyban a membrán két oldalán. Amennyiben az aktuális membránpotenciál megközelíti a Nernst-potenciál értékét, ez azt jelenti, hogy az adott ionra vonatkozóan a kémiai potenciál különbsége (az adott ion koncentrációjának különbsége a belső és külső közegben) és az elektromos potenciál különbsége (membrán-potenciál) egyensúlyt tart egymással. Ilyenkor azt mondjuk, hogy az ion ekvilibriumban van.
ahol Em a membránpotenciál értéke V-ban, R az egyetemes gázállandó, T az abszolút hőmérséklet, F a Faraday-állandó, [ ]e és [ ]i az adott ion koncentrációja a membránnal elválasztott külső (extracelluláris), illetve belső (intracelluláris) térben, P a membránnak az adott ionra vonatkozó permeabilitási állandója. A membránpotenciál aktuális értéke és az adott ionra vonatkozó Nernst-potenciál közötti mindenkori eltérés fontos információt ad arra vonatkozóan, hogy az adott ion mennyire van egyensúlyban a membrán két oldalán. Amennyiben az aktuális membránpotenciál megközelíti a Nernst-potenciál értékét, ez azt jelenti, hogy az adott ionra vonatkozóan a kémiai potenciál különbsége (az adott ion koncentrációjának különbsége a belső és külső közegben) és az elektromos potenciál különbsége (membrán-potenciál) egyensúlyt tart egymással. Ilyenkor azt mondjuk, hogy az ion ekvilibriumban van.
A membránpotenciál kialakításában egy további fontos tényező a membrán féligáteresztő karaktere. Vannak olyan intracelluláris makromolekulák (fehérjék), amelyek méretük alapján képtelenek átdiffundálni a biológiai membránokon, velük szemben tehát a membrán impermeábilis. Ez azért fontos, mert a fehérjék többsége a citoplazmában negatív töltést vagy töltéseket hordozó anionként fordul elő a H+ ion-koncentráció (pH) fiziológiás értékei esetén. A belső térben felhalmozódó és onnan kilépni törekvő fehérje-anionok miatt a membrán belső (citoplamikus) felszíne közelében egy negatív töltésű réteg alakul ki, a membrán külső felszíne közelében pedig a pozitív ellenionok egy pozitív töltésű réteget hoznak létre, ami elektromos potenciál-különbséget (feszültséget) generál. Ezt a feszültséget első leírója nyomán Donnan-potenciálnak nevezzük.
A membránpotenciál negyedik lényeges összetevője az elektrogén ionpumpák működése. Ezek olyan ion-transzport mechanizmusok, amelyek működésük során nem ugyanolyan mennyiségű iont pumpálnak a membránon keresztül az egyik irányban, mint a másikban. Ennek eredményeként eltérő számú töltés kerül a membrán egyik oldaláról a másikra, ami elektromos feszültséget generál. Erre utal az „elektrogén” kifejezés. Egyaránt elektrogén mechanizmus az állati sejtek Na+/K+-ATPáza, valamint a növénysejtek H+-ATPáza. Előbbi egyszeri „körülfordulás” során 3 Na+ iont pumpál az extracelluláris térbe, viszont csak 2 K+ iont vesz fel és juttat az intracelluláris térbe. A H+-ATPáz az intracelluláris térből pumpál H+ ionokat a sejten kívüli közegbe. Mindkét mechanizmus működése során pozitív töltések kerülnek az extracelluláris térbe, azaz az intracelluláris közegben relatíve csökken a pozitív töltések száma. Ez növeli a már amúgy is fennálló negatív túlsúlyt, tehát a membránpotenciál növekedését (hiperpolarizáció) váltja ki.
A nyugalmi membránpotenciált alapértelmezésben a sejtmembrán (plazmamembrán, plazmalemma) által elválasztott térrészre vonatkoztatjuk, tehát a sejt belső és külső környezete között értelmezzük. Értelemszerű emiatt, hogy a membránpotenciál megléte minden élő sejt alapvető sajátossága, és mindaddig fennáll, amíg a sejt az alapvető sejtélettani funkcióit ellátni képes. A membránpotenciál megszűnése az életműködések megszűnését jelenti és a sejtek életképességének tartós károsodását jelzi. Membránpotenciál ezen túlmenően gyakorlatilag minden, membránnal elválasztott kompartment között kialakul, és ennek jelentős szerepe van az egyes kompartmentek közötti transzportfolyamatok szabályozásában. Így pl. jól definiálható membránpotenciál van jelen a mitokondrium belső membránja két oldalán, a kloroplasztisz burkolómembránja (angolul: envelope) két oldalán, valamint a kloroplasztisz tilakoidmembránjai által elhatárolt térrészek között. Ezeknek igen fontos szerepük van az ATP-szintézis folyamatában, hiszen a H+ ionok elektrokémiai potenciál-grádiensének kialakításában a koncentráció-különbség mellett a membránpotenciál az egyik meghatározó tényező.
A sejtmembrán nyugalmi membránpotenciáljának aktuális értéke még állati sejtekben is meglehetősen nagy változatosságot mutat az egyes sejttípusok között, általában legnagyobb (-90 mV) az ideg- és izomsejtekben (az ún. ingerelhető sejtekben, azaz azokban, melyek működése közvetlenül az elektromos tevékenységhez kötött). Ennél általában jóval alacsonyabb a többi sejttípusban, de még a vérben szabadon úszó fehérvérsejtek membránpotenciálja is eléri a -30 mV-ot. A növénysejtek egyik figyelemreméltó tulajdonsága, hogy nyugalmi membránpotenciáljuk általában jóval magasabb még az ingerelhető állati sejtekénél is, és elérheti a -200, sőt helyenként a -250 mV-ot is. A növénysejtek kompartmentációjából adódó másik fontos sajátosság, hogy a citoplazma-vakuolum határán levő tonoplaszt két oldalán is fennáll egy nyugalmi membránpotenciál, amely polaritását tekintve a vakuolumban pozitív.
Az állati sejtekben, különösen az ingerelhető sejtekben jól ismert elektromos jelenség az akciós potenciál, amely az ingerület tovaterjedésének elektromos mechanizmusa. Ma már kiderült, hogy az akciós potenciál hátterében feszültség-érzékeny ioncsatornák működése áll, nevezetesen a depolarizációs szakaszban feszültség-érzékeny Na+-csatornák nyílnak, míg a repolarizációs fázisban feszültség-érzékeny K+-csatornák gondoskodnak a membránpotenciál helyreállásáról. Figyelemreméltó, hogy bár a növények nem rendelkeznek kiegyénült idegrendszerrel, sőt még elszórt idegsejtekkel sem, egyes növényekben mégis megfigyelhető az akciós potenciál. Lefutását tekintve meglepően gyors folyamatról van szó, amely, ha nem is mérhető össze az állati idegsejtekben végigfutó elektromos változások sebességével, mégis az egyik leggyorsabb növényi sejtválasznak tekinthető. A növénysejtek akciós potenciálja során azonban más ionikus mechanizmusok működnek: a depolarizációs szakaszban Cl--csatornák nyílnak és Cl--ionok kiáramlása következik be a citoplazmából az extracelluláris térbe, a repolarizációs szakaszban ugyanúgy, mint az állati sejtek esetén, feszültség-érzékeny K+-csatornák gondoskodnak a membránpotenciál helyreállásáról.